
 |
| A juvenile Edmontosaurus disappears into the enormous, lipped and gummy mouth of Tyrannosaurus. Those of us in the palaeoart community are used to seeing lips on dinosaurs now, but neither the lipped or lipless hypothesis has been given a thorough seeing-to in peer-reviewed literature yet. Until, that is, today. This is the PR art for our new paper, Cullen et al. 2023, that dives into the question of lips for theropod dinosaurs. |
But for all of this online visibility, the question of dinosaur lips and cheeks has received only a little attention from dinosaur researchers. A number of conference abstracts have been presented in this area, but only a handful of these studies have been pushed through peer review to become fully published scientific papers (e.g. Galton 1973; Ford 1997; Knoll 2008; Keillor 2013; Nabavizadeh 2020). It’s largely been dinosaur artists, mostly writing for blogs (such as this, this and this), social media posts or specialist books and magazines (e.g. Witton 2018; Paul 2019), that have provided the bulk of recent conversation on this issue. Whatever merits these discussions have (and there are some very fine, commendable assessments out there), the lack of detailed, authoritative scientific studies has allowed trends in dinosaur facial reconstruction to be shaped by popular culture, palaeoart memes and the opinions of influential palaeoartists more than conventional science. This means that, however comfortable we are with our opinions on dinosaur mouth appearance, this question would benefit from more study, more data and more insight from experts in reptile facial anatomy.
 |
| A visual review of where we are with restoring theropod mouths, from Cullen et al. 2023. Do you prefer your Tyrannosaurus without lips (B and C), or with lips (D and E)? And far more importantly, which of these is better supported by fossil data? |
To that end, today is a good day. I’m part of an international team of researchers publishing a major new paper in Science dedicated to the question of whether theropod dinosaurs possessed lips. Led by Thomas M. Cullen and receiving contributions from Derek W. Larson, Diane Scott, Tea Maho, Kirstin S. Brink, David C. Evans and Robert Reisz, this is the end product of a long-running investigation into theropod faces that was first initiated by Robert 11 years ago. I was invited onto the project at the end of 2020 to create some artwork of lipped and lipless Tyrannosaurus (above) and, once onboard, I’m pleased to say I was able to help contextualise and interpret our data alongside providing some pretty pictures. But the real hard graft of the research was performed by others on the authorship team, so they deserve full credit for the nitty-gritty science and methodological concepts. They also taught me a lot about reptile jaws, teeth and oral soft tissues, so I’m in their debt for this experience.
As you’ll have guessed from the PR artwork that greeted you above, we conclude that yes, theropods almost certainly had lips. As evidence of this, we present multiple lines of evidence that all point to lizard-like scaly tissues covering predatory dinosaur teeth, and perhaps even other types of lizard-like oral tissues as well. Our work can be divided into four independent investigations that collectively support our assertions.
Jaw and tooth form
Firstly, we make some broad-brush comparisons between the tooth orientation and jaw bone morphology of lizards, crocodylians and theropods, some of which will be familiar to those who’ve followed the Lips Debate so far. We note that theropods, early croc-line archosaurs and lizards are similar in having low numbers of jaw bone foramina* distributed along their oral margins, as well as vertically-aligned teeth. Extant crocodylians, in contrast, have splaying teeth and hundreds of evenly-distributed foramina across their skull bones, the functions of which are more specialised than the lip-nourishing jaw openings of lizards.
*If you're new to all this, foramina are small holes in bones that typically house nerve tissues or blood vessels, but can also record other structures, like outgrowths of air sacs.
 |
| Reptile jawbone surface textures and foramina distribution compared, from Cullen et al. (2023). I think this image speaks for itself: the arrangement of theropod jaw foramina (those holes along the jawline) is far more lizard-like than croc-like. |
Some of these observations are not novel as the significance of jaw bone foramina to the theropod lip question has long been recognised (e.g. Bakker 1986; Morhardt 2009; Keilor 2013; Barker et al. 2017; Carr et al. 2017). Our resurrection of this point is, in part, a response to Thomas Carr and colleagues' 2017 paper on Daspletosaurus horneri, which favourably compared tyrannosaur jaw surfaces to those of crocodylians. We don’t think they’re actually much alike at all, especially in foramina distribution, so disagree with that assessment. Carr et al. (2017) also assumed that the thick, immobile facial anatomy of living archosaurs — crocs and birds — was ancestral to their entire group, including non-bird theropods. We question this too. Like theropods, the jaw surface properties and tooth orientations of early croc-line archosaurs recall those of lizards more than modern crocodylians, probably reflecting a different soft-tissue configuration. We thus agree with the increasingly evidenced view that the faces of living archosaurs are specialisations suited to very particular lifestyles and that the dinosaur extant phylogenetic bracket is of limited use for inferring their facial anatomy.
Enamel, hydration, and tooth wear
Secondly, we discuss the damage and wear inflicted on permanently exposed teeth, a conversation that is mostly about enamel hydration. Enamel is one of the hardest tissues that animals can synthesise and is thus highly resistant to damage, but its resilience is dependent on moisture. Hydrated enamel is more plastic, and thus more resistant to abrasion, than dehydrated enamel, which is brittle and prone to cracking and breaking under strain. To that end, teeth emerging from oral margins tend to be more damaged and worn than those kept within a moist, sealed mouth. This difference can be seen with the naked eye but is particularly obvious under microscopic examination. In our paper, we show that the tips of alligator teeth are shorn off on their outward-facing, exposed surfaces, with both the enamel and several layers of dentine worn to a flattened edge. Tooth dehydration almost certainly factors into crocodylians frequently suffering from broken and cracked dentition, and they have to replace their teeth regularly (something like 45-50 times in a lifetime — Grigg and Kirchner 2015) to maintain a set of fully functional jaws.
These observations give us a clear hypothesis regarding theropod oral tissues. If they were permanently exposed, theropod teeth should show, at minimum, similarly obliterated enamel and dentine layers at their tips. The poster children of exposed dinosaur teeth, adult tyrannosaurids, are especially relevant here as they replaced their teeth at a very slow, sometimes even biennial rate (Erickson 1996). What’s more, they engaged in particularly violent, tooth-on-bone feeding strategies. So, if any theropods are going to have knackered, abraded teeth, it’s tyrannosaurids.
 |
| Detailed comparisons of tyrannosaurid (upper row) and crocodylian (lower row) tooth wear. Note how the Daspletosaurus tooth, despite being over 500 days old, is intact despite dinosaurs teeth having particularly thin enamel layers. The alligator tooth tip, by contrast, has not only lost the enamel coating on its outer surface, but also several layers of underlying dentine. A fully intact, enamel-covered erupting alligator tooth is shown in panel H to show that these are features of wear, not the original tooth condition. From Cullen et al. (2023). |
But extracting a fully grown, c. 510-day-old tooth from a Daspletosaurus maxilla revealed a tooth tip in great condition. Both the inner and outer surface retained their relatively thin enamel covering and the only minor damage found was on its medial, inward-facing surface, possibly as a result of occasional tooth-on-tooth contact. This confirms what has generally been observed for theropod dental wear in other studies and conflicts with what we'd expect from a dehydrated, perpetually exposed tooth. We take this as evidence of theropod dentition being maintained in a moist, well-hydrated setting, and being located within a lipped mouth is realistically the only way this might be achieved.
Too big to sheath?
We also address the possibility that some theropods had teeth that were simply too big to cover with lips (e.g. Ford 1997). Our focus here is not on the widely known, but still surprisingly prevalent issue of artists and researchers not accounting for tooth slippage in fossil specimens**. Rather, we focus on realistic estimates of tooth crown height when they are fully socketed in theropod mouths. We calculated a ratio of tooth crown height to skull length for 37 theropod specimens and compared them with the same metric in 40 varanids, the monitor lizards. Varanids are, of course, well-known for possessing large, theropod-like teeth, as well as copious amounts of lip and gum tissue.
**This is the decay of tooth-anchoring ligaments resulting in teeth sliding somewhat from their sockets, preserving them at longer lengths than they held in life.
 |
| Tooth size: theropods vs varanids. It turns out that both groups have similarly-sized teeth relative to skull length and, while this doesn't directly tell us if theropods had lips, it shows that their dentition was of a size that we know can be sheathed by extra-oral tissues today. From Cullen et al. (2023). |
Plotting these data showed that theropod and monitor teeth are about the same size for their skull lengths and even increase in proportion at the same approximate rate. But the winner of the biggest tooth contest wasn’t something like T. rex: it was the varanids. Some monitor species, like the crocodile monitor Varanus salvadorii, have almost cartoonishly-large dentition. From this, we suggest that theropods did not need unprecedentedly big lips to cover their mouths and the largest theropods wouldn't look, in terms of lip proportions, very different to something like a komodo dragon. Indeed, we note that monitors are able to cover their teeth with the same basic configuration of labial and gingival tissues across a 12-fold size difference. The discrepancy between the largest monitor skull and our largest theropods is only half that: 6-fold. So if lizard lips and gums can scale 12 times over without substantial anatomical deviation, perhaps they could stretch to cover the teeth of much bigger animals without much change, too? Whether we've realised it or not, a lot of us already evidently believe this is possible, given the abundance of lippy, monitor-esque mosasaur reconstructions.
Sealing the deal
These are the points we cover in the main paper but, this being an article in Science, it’s essential to also check out the supplementary files for additional discussion and context. Therein we raise another point that resulted from our efforts at reconstructing a scientifically-informed illustration of a lipless Tyrannosaurus: it’s really, really hard, maybe even impossible, to seal at least some theropod mouths without lips. Forming an oral seal, even if it’s just by pressing lipless jaws against one another, is important to avoiding dehydration as well as maintaining basic oral health and hygiene. We could not, however, find a way to reconstruct T. rex jaws without leaving a gap behind their maxillary teeth. I actually pushed our lipless reconstruction (Fig. 1B in the paper) a little beyond what I think is reasonable and we're still left with a small opening.
We are not the first people to ponder this issue, and dinosaur literature has contrasting views on how far theropods could close their jaws. Some authors propose that theropod mandibles could be pulled way up into the cavity of the upper jaw and have even identified landmarks for the resting position of the lower teeth (below). These include depressions in the walls and roof of the upper oral chamber that seem suited to act as socket-like structures for receipt of the lower dentition (e.g. Molnar 1991; Ford 1997; Currie 2003; Hendrickx et al. 2014). Others, most notably Tyler Keillor (2013) in his excellent book chapter on restoring the face of the “Jane” Tyrannosaurus, have questioned this idea on grounds that theropod mandibles can’t close so tightly without literally bashing into problems.
 |
| Examples of landmarks suggested to record the resting poses of theropod lower jaws. Maxillary wall sockets are depressions in the internal wall of theropod oral cavities, and some theropods are also preserved with round depressions in the roofs of their mouths. Neither are universal features of all theropods, however. Images from Osborn (1912), Lü et al. (2014) and Cullen et al. (2023). |
Reconstructing the face of T. rex for our paper saw us agreeing with Tyler's conclusions. At a certain point of mouth closure, theropod lower jaws collide with bones under the eye socket (specifically, the ectopterygoid) so that further adduction either requires the jaws to literally crush themselves shut, or else the bones of the posterior skull act as a hinge, swinging the jaw tip into the mouth but dislocating the jaw joint. Ford (1997) proposed that a notch in the ectopterygoid accommodated the closed lower jaw during mouth closure but we don’t think this is plausible. Theropod ectopterygoids can be complex shapes and yes, some have regions that superficially look like they could nestle the lower jaw, but these were almost certainly filled by deep jaw muscles in life (e.g. Gignac and Erickson 2017). There are, of course, theropod skulls preserved with their jaws tight shut in the fossil record but we have to be careful assuming these represent in vivo conditions, given how routine processes of decay and fossilisation can pull and crush carcasses into unnatural configurations.
Inspired by this, we devote some discussion to how theropods posed their closed jaws in life. Beyond ruling out impossible, jaw-busting configurations, it's difficult to know exactly how tightly theropods held their mouths, but this is something for artists to consider. X-rays and scans of lizard carcasses show that their jaws are far from clenched shut when their mouths are closed and, in some species, their upper and lower dentition barely overlaps. If we go "full lizard" with our theropod reconstructions, where we apply minimal overlap of the upper and lower toothrows, their lips would have been deep and their snouts much taller than we’re used to. We play about a little with this visually in the paper and I was struck at the blocky, chunky cranial profile of our lizard-like, loose-mouthed T. rex (below).
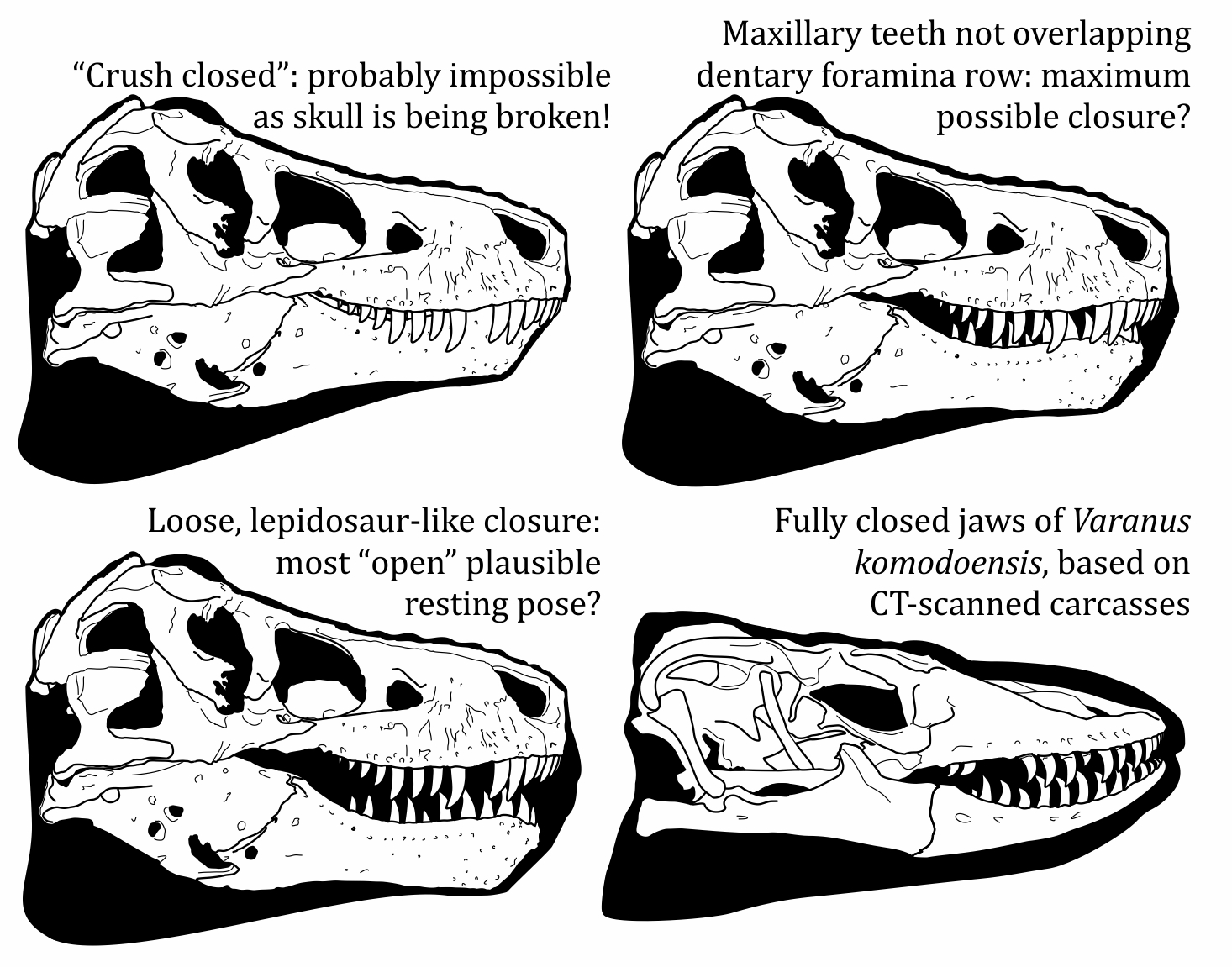 |
| Some of our experimentation with theropod mouth postures. The "crush closed" pose is almost certainly impossible, but it's hard to say how relaxed theropod resting gapes may have been held. If modern lipped reptiles are anything to go by, they may have been held far more "open" than we're used to. Modified from Cullen et al. (2023). |
Conclusion: theropod jaws only make sense if they had lips
Putting all this together, our investigations of jaw structure, tooth size, tooth wear and jaw closing all point to the same inference: theropod jaws don’t make much anatomical or functional sense without lips of some kind. To validate the alternative lipless model, we have to engage in a lot of special pleading and scientific weaselling. Exposed theropod teeth would have to be unprecedentedly resistant to wear; all our understanding of jaw structure and foramina distribution correlating with oral soft tissues would have to be wrong, and theropods would need to be unique in not bothering to create oral seals. If we're being good scientists, we can’t currently say that theropods definitely had lips, drop the microphone and walk off stage, but I think we've made it far more challenging for anyone to legitimately object to the lipped theropod hypothesis. Time will tell on that front.
In addition to substantiating the lipped hypothesis of theropod appearance, our hope is that our paper may establish some lines of inquiry for the oral tissues of other extinct animals. Many of the most extreme dentitions to ever evolve belong to fossil taxa, after all, and theropods are far from the only species with uncertain facial appearances. What of nimravids, gorgonopsians, uintatheres, or Thylacosmilus? And what, for that matter, of the superficially crocodile-like spinosaurids and other weird theropods — were they lipped or not? If our ideas hold water, they provide a relatively straightforward way of deducing whether the teeth of these animals were held within oral tissues.
We can't, of course, finish without some brief notes on the life appearance of lipped theropods. We address this a little in our supplementary information and conclude that lepidosaurs, the lizards and tuataras, are the best modern analogue for theropod lips and gums. This is, admittedly, a “best of a bad situation” recommendation because there are plenty of differences between theropod and lepidosaur jaws that preclude total confidence in their comparison, but we only have so many extant reptile groups to choose from and lepidosaurs are, on the whole, morphologically closer to theropods in areas we think are influential on labial and gingival tissues.
 |
| What did lipped theropods actually look like? Lizard mouths have a lot more soft-tissue surrounding their teeth (jaw cross sections show a komodo dragon, B, and alligator, C), and this might be something we need to bring into our theropod artwork. The gaping T. rex shown here is outfitted not only with large lips, but also a conservative 25% of its tooth height covered with gums. From Cullen et al. (2023, supplementary data). |
This being the case, our model for theropod mouths is that they were sealed by non-muscular*** lips covered with scales or — to hedge our bets a little more — whatever epidermal covering was present on the side of the snout. Lepidosaurs show variation in lip size, with most having generous upper lips but some having thinner lower lips than others. This variation continues to their gums. Lepdiosaur gingivae are more voluminous than those of mammals and crocodylians and generally cover at least 20-25% or so of tooth crown height. This is why lizard teeth aren’t always that conspicuous in their open mouths. Varanids take these enlarged gingivae to an extreme, hiding almost all of their formidable teeth with enormous gums. We currently don’t have much insight into where theropods sat within this range. Paul (2019), independently of our study, advocates for full monitor-like conditions for theropods, and this might be possible, but we can't rule out smaller gums or, indeed, a unique theropodan take on oral soft-tissues at this time. We propose, however, that since extant lipped reptiles have at least 25% of their tooth crown heights covered with gums, we should apply that to theropods, too. We’ve explored this in our paper and PR art with that gummy, lippy T. rex shown above.
***It’s not strictly true that lizards have no muscles around their mouths. Some agamids have muscles that move their lips or flaps of mouth-adjacent skin for communicative purposes. I'm no expert on these structures, but I think it’s fair to assume that they are specialisations of their respective lineages, not the remnant of a once ubiquitous, clade-wide lepidosaur ability.
And I think we’ll leave it there for now. There’s more to say on the tyrannosaur restorations we created for the paper with their small eyes and some of their skin details, but at least some of that discussion needs to wait for developments in other research I’m involved in (I'm so, so sorry, Dave). For now, I’ll thank my coauthors once again for inviting me onto such a great project, and I’ll leave you with this handy infographic summary of our research, which you can share around the internet to instigate discussion of dinosaur life appearance wherever you may be. The QR code in the corner will take you directly to the paper so, if you or anyone you encounter wants to know exactly what we have to say about theropod lips, you can always find it from this image.

Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored through Patreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusive Patreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
- Bakker, R. T. (1986). The dinosaur heresies: new theories unlocking the mystery of the dinosaurs and their extinction. William Morrow.
- Barker, C. T., Naish, D., Newham, E., Katsamenis, O. L., & Dyke, G. (2017). Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii. Scientific Reports, 7(1), 3749.
- Carr, T. D., Varricchio, D. J., Sedlmayr, J. C., Roberts, E. M., & Moore, J. R. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7(1), 1-11.
- Cullen, T. M., Larson, D. W., Witton, M. P., Scott, D. Maho, T. Brink, K. S., Evans, D. C. and Reisz, R. (2023). Theropod dinosaur facial reconstruction and the importance of soft tissues in paleobiology. Science, 379, 1348-1352.
- Currie, P. J. (2003). Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada.
- Ford, T. L., 1997, Did Theropods have Lizard Lips? Southwest Paleontological Symposium – Proceedings, 1997, 65-78.
- Galton, P. M. (1973). The cheeks of ornithischian dinosaurs. Lethaia, 6(1), 67-89.
- Gignac, P. M., & Erickson, G. M. (2017). The biomechanics behind extreme osteophagy in Tyrannosaurus rex. Scientific Reports, 7(1), 2012.
- Grigg, G., & Kirshner, D. (2015). Biology and Evolution of Crocodylians. Csiro Publishing.
- Hendrickx, C., & Mateus, O. (2014). Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in nonavian theropods. PloS one, 9(3), e88905.
- Keillor, T. M. (2013). Jane, in the flesh. In: J. M. Parrish, R. E. Molnar, P. J. Currie, E. B. Koppelhus, (eds.). Tyrannosaurid Paleobiology. Indiana University Press.
- Knoll, F. (2008). Buccal soft anatomy in Lesothosaurus (Dinosauria: Ornithischia). Neues Jahrbuch fur Geologie und Palaontologie-Abhandlungen, 248(3), 355-364.
- Lü, J., Yi, L., Brusatte, S. L., Yang, L., Li, H., & Chen, L. (2014). A new clade of Asian Late Cretaceous long-snouted tyrannosaurids. Nature communications, 5(1), 3788.
- Molnar, R. E. (1991). The cranial morpholgy of Tyrannosaurus rex. Palaeontographica. Abteilung A, Paläozoologie, Stratigraphie, 217, 137-176.
- Morhardt, A. C. (2009). Dinosaur smiles: Do the texture and morphology of the premaxilla, maxilla, and dentary bones of sauropsids provide osteological correlates for inferring extra-oral structures reliably in dinosaurs?. Western Illinois University.
- Nabavizadeh, A. (2020). New reconstruction of cranial musculature in ornithischian dinosaurs: implications for feeding mechanisms and buccal anatomy. The Anatomical Record, 303(2), 347-362.
- Osborn, H. F. (1912). Crania of Tyrannosaurus and Allosaurus; Integument of the iguanodont dinosaur Trachodon. Memoirs of the AMNH; new ser., v. 1, pt. 1-2.
- Paul, G. S. (2019). Non-ornithischian dinosaurs probably had lips. Here’s why. Prehistoric Times 127, 44-49.
- Witton, M. P. (2018). The Palaeoartists' Handbook. Crowood Press.



.jpg)









