
 |
| Juvenile, subadult and a big, old adult Rhamphorhynchus muensteri forage in a Jurassic lagoon. We know more about the position of this pterosaur in Mesozoic food webs than any other thanks to its excellent fossil record. The floating posture here is based on data from Hone and Henderson (2014). |
*We see these restored for numerous long tailed pterosaurs, but only Rhamphorhynchus is known, for fact, to have this 'classic' tail vane morphology.
Snap my fish up
What did Rhamphorhynchus eat? At least three specimens suggest that it foraged for fish of various sizes. The most famous example of such a fossil is sometimes referred to as the 'greedy guts Rhamphorhynchus', an animal which swallowed a fish almost as long as its own torso. This specimen represents a smallish individual, first described by Wellnhofer (1975), with a partly articulated, partly digested fish fossil preserved inside the posterior 60% of the rib cage (below). The orientation of the fish tail fin suggests the fish was swallowed head first, and we have to assume that some major distension of the throat occurred when doing so. That pterosaurs were capable of doing this isn't that surprising considering that extant pterosaur relatives - crocodylians and birds - have mobile throats which aid swallowing of big food items. The relative proportions of these gut contents suggests this animal must've bolted its food like these modern archosaurs, a behaviour rare among mammals but known to occur in at least one large, yellow, bipedal primate. This specimen also presents several elongate elements which defy easy explanation: they're suggested as other food items or even bits of preserved gut tissue by various authors (dark grey in the line drawing below). |
| The fossil, interpretive drawing, and restored reality of Jurassic table manners. Disgraceful. Line drawing after Wellnhofer (1975). |
The one, the only, pterosaur coprolite
Of the many questions that keep pterosaur experts up at night - what are they related to? how did they work as functional organisms? how are they related to each other? - none has been greater than what their poop was like. Expelled waste is a common form of fossils in some localities, but remained entirely elusive for flying reptiles until last year when Dave Hone and colleagues (2015) identified the first pterosaur coprolite dropping out of a complete Rhamphorhynchus specimen. Finally, pterosaur workers can sleep easy.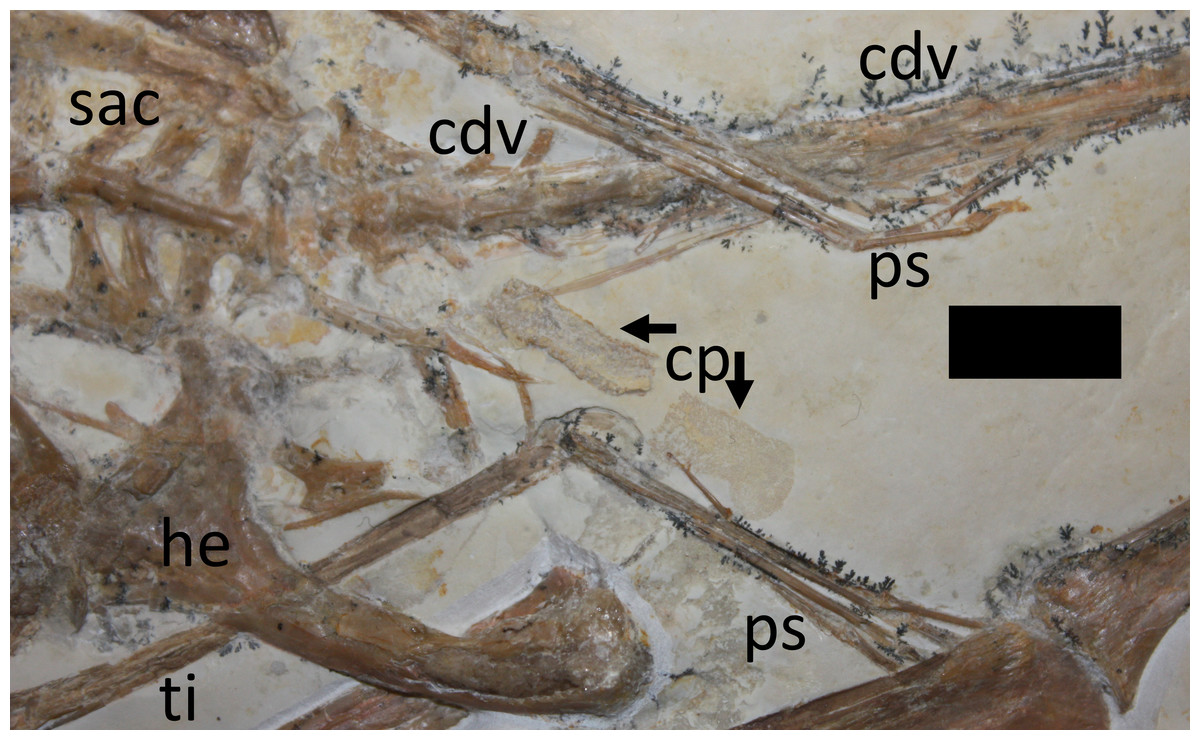 |
| Rhamphorhynchus muensteri specimen with coprolite (cp). From Hone et al. (2015). |
Fossilised food chains
Several Rhamphorhynchus specimens reveal it was prey to other Jurassic animals. Among the least commonly discussed is a small pellet produced by something like a fish or crocodylomorph which contains several Rhamphorhynchus wing bones (Schweigert et al. 2001). This is a rare example of Rhamphorhynchus from the Nusplingen Limestone, a unit lithologically similar to Solnhofen but slightly older. As is so often the case with such fossils, the identity of the pellet maker remains elusive. My own suspicions are of a piscine origin, as modern crocodylians don't tend to spit out bones (they digest them, only regurgitating hair, feathers and other keratinous tissues that are difficult to break down). That may not have been true for fossil crocs, of course. |
| Rhamphorhynchus vs. Aspidorhynchus. I guess we should call this one a draw? From Frey and Tischlinger (2012). |
Among the most remarkable Rhamphorhynchus fossils are five instances where it is preserved alongside the predatory fish Aspidorhynchus acutirostris, a long-bodied species that is often much larger than its Rhamphorhynhus prey (Frey and Tischlinger 2012; Weber 2013). These fossils are palaeoecologically notable for three reasons. Firstly, large Solnhofen vertebrates are hardly ever associated, and yet we have five instances of this same pterosaur and fish species being found in touching, or near touching, proximity. Secondly, all five are exquisitely preserved - one example includes a 'mummified' pterosaur with wing membranes, and all are completely articulated. Thirdly, the Rhamphorhynchus are invariably positioned around the skull of the fish, as if a specific, repeated behaviour saw these animals preserved together. These particulars make chance association of these animals unlikely and imply Aspidorhynchus sought out Rhamphorhynchus as food, probably hunting live specimens rather than scavenging floating corpses (as indicated by the excellent preservation of the individual pterosaurs). Frey and Tischlinger (2012) provide a plausible scenario for the deaths of these animals: they reason the Aspidorhynchus tackled prey that became entangled in their jaws before accidentally entering the toxic bottom waters of the Solnhofen waterways. In these anoxic depths the tangled pair would die pretty quickly (if the pterosaur wasn't already dead, of course), leaving us with perfectly preserved bungled predatory acts. The icing on the cake of these specimens is that one of these Rhamphorhynchus specimens has already been mentioned here - that individual with a fish in its throat (Frey and Tischlinger 2012). In this specimen at least, we can assume the pterosaur ate a fish shortly before being attacked itself: a rare instance of a fossil food chain.
The bigger picture of Rhamphorhynchus palaeoecology
Collectively, we have 10 specimens of Rhamphorhynchus telling us something about its position in Mesozoic food webs. That's not bad going for a pterosaur, a famously rare type of fossil, and is actually a pretty good palaeoecological record for any fossil vertebrate. 10 specimens is not enough to tell us everything about the lifestyle of a fossil animal, but does allow us to paint a general picture. They show us that Rhamphorhynchus was adapted to foraging on pelagic prey - often small, probably live fish - and that it must have spent a good amount of time in or around water, as it was clearly an attractant to aquatic predators. These specimens gel neatly with general models of Rhamphorhynchus lifestyle interpreted from their functional anatomy. It's generally thought that Rhamphorhynchus was adapted for life along shorelines - basically a Mesozoic gull. It's not uncommon to be suspicious of such claims nowadays, it being realised that the 'Mesozoic seabird equivalent' almost became a trope, or at least a tremendous over-generalisation, of pre-21st century pterosaur science. But in this case, a gull-like lifestyle is a cogent hypothesis based on studies of wing shape, flight style, tooth and jaw apparatus, and limb function (e.g. Wellnhofer 1975; Hazlehurst and Rayner 1992; Witton 2008, 2015; Ösi 2011). Perhaps research on Rhamphorhynchus was involved in creating the stereotype of pterosaurs as Mesozoic seabirds, but we should not regard it as a victims of this stereotyping itself.But we should ask ourselves why Rhamphorhynchus palaeoecology is comparably well-represented in the fossil record. It might be something as simple as preservational conditions, but there are plenty of pterosaur Lagerstätten, some of them containing close relatives of Rhamphorhynchus in reasonable abundance, which provide no information about dietary preferences or interactions with predatory species. Is there something intrinsic to Rhamphorhynchus which makes it special? I find several reasons to wonder if Rhamphorhynchus was an atypically aquatic species, not only flying and feeding above water but actually routinely entering it. Firstly, Rhamphorhynchus has a hatchet-shaped deltopectoral crest (the process on the humerus which anchors the flight muscles), a small (or at least narrow) torso and short legs: these are features which Habib and Cunningham (2010) link to routine and efficient aquatic takeoff. Secondly, tests of pterosaur swimming suggest that Rhamphorhynchus had a pretty stable floating posture - not something that can be said for all pterosaurs (Hone and Henderson 2014). Thirdly, it also has comparably large but delicately constructed feet, which might suit paddling, as well as a slender set of forelimb bones (Witton 2015) which recall the streamlined arm bones of swimming and diving birds (Habib and Ruff 2008). Prolonged bouts of swimming might also account for its general abundance, excellent preservation and all-round good fossil record: being immersed in water puts it a step closer to being preserved than other, less aquatic species. Who knows: perhaps it even dived into Solnhofen's toxic bottom waters on occasion, explaining why so many specimens are excellently preserved? Hmmm.... perhaps this warrants further investigation.
Coming soon: could dinosaurs - gasp - lie down on their sides? My take on the greatest of palaeoart debates.
Rhamphorhynchus is supported by fish; this blog is supported by Patreon
The paintings and words featured here are sponsored by the finest human beings on the planet, those folks who support me at Patreon. Backing my blog for as little as $1 a month helps me churn out researched and detailed articles and paintings to accompany them, and in return you get access to bonus blog content: additional commentary, in-progress views and high-resolution artwork, and even free prints. Accompanying this post, we're going to look at the bigger picture of pterosaur palaeoecology: azhdarchids and Rhamphorhynchus are just two lineages with palaeoecological records - what about the rest of Pterosauria? Sign up to Patreon to join that discussion!References
- Frey, E., & Tischlinger, H. (2012). The Late Jurassic pterosaur Rhamphorhynchus, a frequent victim of the ganoid fish Aspidorhynchus?. PloS one, 7(3), e31945.
- Habib, M. & Cunningham, J. 2010. Capacity for Water Launch in Anhanguera and Quetzalcoatlus. Acta Geoscientica Sinica. 31, 24-25
- Habib, M. B., & Ruff, C. B. (2008). The effects of locomotion on the structural characteristics of avian limb bones. Zoological Journal of the Linnean Society, 153(3), 601-624.
- Hazlehurst, G. A., & Rayner, J. M. (1992). Flight characteristics of Triassic and Jurassic Pterosauria: an appraisal based on wing shape. Paleobiology, 18(04), 447-463.
- Hone, D. W., Habib, M. B., & Lamanna, M. C. (2013). An annotated and illustrated catalogue of Solnhofen (Upper Jurassic, Germany) pterosaur specimens at Carnegie Museum of Natural History. Annals of Carnegie Museum, 82(2), 165-191.
- Hone, D. W., & Henderson, D. M. (2014). The posture of floating pterosaurs: Ecological implications for inhabiting marine and freshwater habitats. Palaeogeography, Palaeoclimatology, Palaeoecology, 394, 89-98.
- Hone, D., Henderson, D. M., Therrien, F., & Habib, M. B. (2015). A specimen of Rhamphorhynchus with soft tissue preservation, stomach contents and a putative coprolite. PeerJ, 3, e1191.
- Ősi, A. (2011). Feeding‐related characters in basal pterosaurs: implications for jaw mechanism, dental function and diet. Lethaia, 44(2), 136-152.
- Schweigert, G., Dietl, G. & Wild, R. (2001). Miscellanea aus dem Nusplinger Plattenkalk (Ober-Kimmeridgium, Schwäbische Alb) 3. Ein Speiballen mit Flugsaurierresten. Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereines, 83, 357-364
- Weber, F. (2013). Paléoécologie des ptérosaures 3. Les reptiles volants de Solnhofen, Allemagne. Fossiles. 14. 50-59.
- Wellnhofer, P. 1975. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Palaeontoographica A, 149, 1-30.
- Witton, M. P. (2008). A new approach to determining pterosaur body mass and its implications for pterosaur flight. Zitteliana, 28, 143-159.
- Witton, M. P. (2015). Were early pterosaurs inept terrestrial locomotors? PeerJ, 3, e1018.
Oh my god, Mark. That pun in the first header. It's too good. It made my day.
ReplyDeleteYou know, their floating position vaguely reminds me of one of these things:
ReplyDeletehttp://www.swimways.com/images/Product/medium/31.jpg
I remember seeing the Mr. Greedy Guts skeleton in a pterosaur book. They pointed out those little unknown, banana-shaped bits and I feel the need to ask, did we ever find out what they were? My guess was either another food source, undigested bits of fish or some kind of internal parasite.
ReplyDelete