
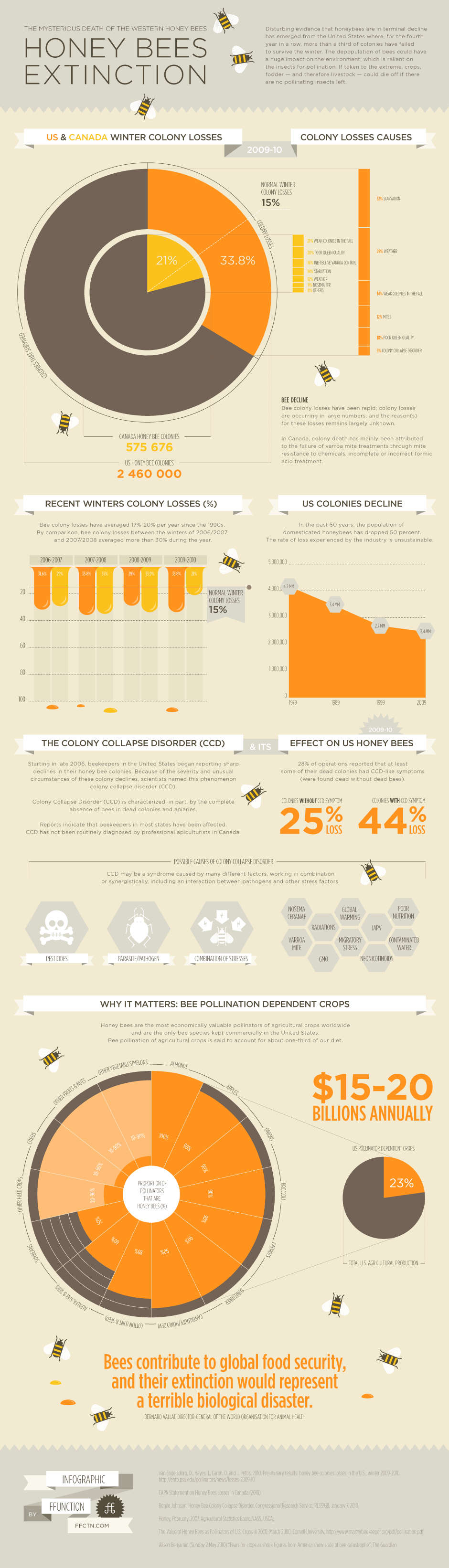
To celebrate the launch of the store, I thought it would be cool to show four of my favourite new pieces of art generated within the last few months. These all represent private commissions which I have permission to post and sell as prints. If you want your own copy, you know where to go...
Dreadnoughtus dwarfs Talenkauen, is happy
 |
| "Oh, you say you're a medium-sized dinosaur? Sorry, it's hard to hear you with my head all the way above the trees here." Experts predict Dreadnoughtus schrani was jerk it was to other, smaller species like the iguanodont Talenkauen santacruensis. Print. |
First up is Chris Wummer's commission of giant, latest Cretaceous titanosaur Dreadnoughtus schrani, an animal which needs little introduction after the publicity of its discovery last year. Dreadnoughtus was publicised as the most massive terrestrial animal of all time at 59 tonnes, but regular readers of the palaeoblogosphere may know that sauropod guru Matt Wedel questioned this over at SVPOW! through rough volumetric estimates of mass and, later, when considering the restored Dreadnoughtus trunk as too long. Palaeoartist Greg Paul has also provided contrary comment on the 59 tonne estimates and restored proportions (although I'm not really sure what context that article is presented in - it looks like an unpublished MS). Estimating the mass of any extinct animal is difficult and especially so at the extreme sizes represented by giant titanosaurs, but there seems good reason to think the Dreadnoughtus holotype individual achieved a mass of 30-40 tonnes. That's still very big of course, but within fairly 'typical' ranges for giant titanosaurs.
There are two versions of the Dreadnoughtus image shown here. Chris wanted the picture to have personal relevance and so asked for his house to be included. That choice was inspired by his residence in Philadelphia, the city were Dreadnoughtus was studied and unveiled to the world. Switching between the version with familiar modern objects and a completely 'natural' scene reinforced how difficult it is to show absolute prehistoric animal size without a frame of reference: Dreadnoughtus looks a lot smaller when its head isn't clearing a rooftop. Two ornithopods - the 4 m long iguanodont Talenkauen santacrucensis - were added to this version to help stress the size of the sauropod. It's still difficult to appreciate a precise size of the sauropod in this image, but hopefully it at least looks very big, which might be the best we can hope for in images without obvious scale references.
Deinonychus pair in the swamps
 |
| Two Deinonychus antirrhopus either taking a moment to drink, or looking at something really interesting at the bottom of that pool. Print. |
Next up is Patrick Murphy's pair of Deinonychus antirrhopus. The Early Cretaceous dromaeosaur Deinonychus has been restored so many times that it's difficult to come at it from a fresh angle. I thought one way to do that was to not show it on open plains, but in a backswamp. Deinonychus is known from two geological units, the Cloverly and Antlers formations, both of which represent sediments deposited by ancient, subtropical rivers and their floods. Some sediments in the Antlers Formation represent large (10 m wide or more) abandoned river channels, complete with evidence of soils, low velocity or still water, and ancient vegetation (Hobday et al. 1981). The depicted animals are meant to have recently eaten something - their muzzles are still read with blood - and popped down to their local swamp for a drink and some shade. I imagine that these guys are set to sit down and digest after this, waiting until they get hungry enough to chase prey again.
The arms of the foreground animal are pressed tight to the body in the manner proposed by palaeoartistic Queen of the maniraptorans, Emily Willoughby, rather than held half-folded as we're more used to seeing them. As Emily explains, there is good reason to think the 'arms out' postures we're used to is nonsensical - animals just don't carry themselves like that (including ourselves: our arms don't just hang limp - we fold, stow and hold them when they're not in use).
These guys were a lot of fun to paint: Deinonychus has an appealing character - a sort of mash up of a wolf and a raptorial bird - which is fun to try to capture. My thought is that Deinonychus should always look like an animal which we would admire and revere, but would purposely avoid close proximity with.
Parasaurolophus, alone with other dinosaurs
 |
| Parasaurolophus walkeri, wondering where his friends are. Print. |
Delano DuGarm's Parasaurolophus walkeri brings us back to the Late Cretaceous, specifically the Campanian. Delano's brief was for a fairly minimalist scene, which I think matches one part of the 'Campanian story' quite well. By this time some of the fauna and flora we think of as epitomising the Mesozoic were already gone or showing clear evidence of decline, including ichthyosaurs, some dinosaurs, pterosaurs and ammonites. Although some taxa were doing fine in this interval, and even radiating, seeds of change were already being sown for Cretaceous biospheres. We have to wonder how long many 'classic' Mesozoic groups would have lasted even without the global catastrophes occurring at 66 million years ago: even without them, the post-Mesozoic world might have been quite different.
Delano's lone Parasaurolophus painting gave a good opportunity to hint at this changing world. The left of the painting features a few (speculative) wading birds and two bird flocks leaving the trees - these, of course, are the 'new dinosaurs' that will live on through the late Cretaceous troubles. The Parasaurolophus on the right looks a bit big and cumbersome by contrast, sort of like an old design which can't compete with new technologies. Aiding this comparison is the relative chunkiness of the Parasaurolophus skeleton: hadrosaurs are hardly a svelte bunch, but the bones of Parasaurolophus are especially big and robust, with expanded areas for muscle attachment. As far as I'm aware, the significance of this is unknown (but let me know otherwise in a comment below!).
Carnotaurus with a difference
 |
| Azhdarchids > theropods, as demonstrated by this lousy predation attempt by Carnotaurus sasteri. Print. |
Finally, we're popping back to Maastrichtian South America for Chris Tait's Carnotaurus sasteri vs. azhdarchids image. An obvious artistic departure from the rest, this is an attempt to achieve a comic-book style in line with Chris' intention to give this to his son as a present. I've tampered with minimalist, comic-book styles before and quite enjoy it. Comic-book palaeoart - especially Ricardo Delgado's Age of Reptiles graphic novels - has influenced my work since the age of nine because of the energy, character and personality infused into the animals. Of course, you have to try hard not to find character in animals like Carnotaurus which, with its strange proportions and anatomy, looks almost like work of comic book fiction already (must... resist comment... about fictional theropod design and Jurassic World...). Carnotaurus, like other abelisaurs, was adapted for speed more than manoeuvrability, and this attempt to grab a passing pterosaur snack is an example of how nimble, agile prey might easily evade one. The pterosaurs shown here are quite small, which might seem odd for very late Cretaceous azhdarchids - aren't the small pterosaurs meant to be gone by then? Fragments of pterosaur jaw from Late Cretaceous Hungary indicate that some azhdarchid species retained small absolute body sizes even when most of the group represented medium-giant species (Prondvai et al. 2014). The discovery of these smaller Late Cretaceous pterosaurs does not buck the overall trend of average pterosaur size increase throughout the Mesozoic of course, but it does show that there were some exceptions to this wider trend.
Yes yes yes... but how are the bees doing?

Regular readers will know that I'm donating all funds from February sales of one print to the Bumblebee Conservation Trust. The good news is that I'm now up to a donation of £130, and there's still eight days left to get your order in. I'm really happy to have sold enough of these to break £100 - huge thanks to everyone who's bought one - and exceeding £150 is my new goal.
It's now easier than ever to buy a copy of the bee-charity print, so you can get yourself a copy and help our struggling wildlife with just a few mouse clicks. Prices start at £20 (+shipping), and I'm giving as much as I can from each sale to the trust.
References
- Hobday, D. K., Woodruff, CM, Jr., McBride, MW. (1981), Paleotopographic and structural controls on non-marine sedimentation of the Lower Cretaceous Antlers Formation and correlatives, north Texas and southeastern Oklahoma. Recent and ancient nonmarine depositional environments, 71-87.
- Prondvai, E., Bodor, E. R., & Ősi, A. (2014). Does morphology reflect osteohistology-based ontogeny? A case study of Late Cretaceous pterosaur jaw symphyses from Hungary reveals hidden taxonomic diversity. Paleobiology, 40(2), 288-321.




{kind=link}
{kind=link}
{kind=link}