 |
| My mural of a Diplodocus carnegii herd, currently keeping Dippy, the Natural History Museum's Diplodocus cast, company in Dorset County Museum. At 4 x 2 m, it's the third biggest picture I've ever done, and - as positioned at the museum - the most visible. |
If you head to
Dorset County Museum at some point before May 9th 2018 you'll be able to see a genuine dinosaur celebrity: the Natural History Museum's 'Dippy'
Diplodocus skeleton, on the first stint of its
'Dippy On Tour' campaign of UK museums. The trip is well worth the visit even if you're familiar with the specimen from the NHM's Hintze Hall. A mezzanine around the skeleton, and the smaller size of the exhibition space, allows visitors to get closer to Dippy than ever before, and you can see the specimen from elevated positions unavailable at the NHM. If you're a sauropod fan in the UK, this might be your best chance to see this specimen up close and personal. It's free to see the skeleton, but you do need to
book in advance - the tickets are flying off the shelves, so don't expect to just walk in.
 |
| A discerning audience checks out my prints at Naturally Curious. Say, some of those images look a little Life-through-the-Ages II-y... |
Alongside Dippy is a collection of art entitled
Naturally Curious, works by four different artists inspired by fossils and the natural world. My work is among them (above) and includes a 4 m wide mural based on the Dippy specimen and its palaeoenvironment - the same image that welcomed you to the post. It's not placed with the rest of my work but hanging right next to the Dippy skeleton itself - the first time a detailed artistic restoration has been associated with the specimen since the 1980s when a scale model stood next to its tail. This mural, commissioned by the Dorset County Council, was a great opportunity to bring Dippy's visitors up to speed on the latest ideas on sauropod dinosaur life appearance (as well as very flattering for me - it's not every day you're asked to display art next to one of the most famous dinosaurs in the world). The process involved learning a lot about the Dippy specimen, applying some new ideas about dinosaur anatomy to
Diplodocus, and looking into the specifics of Dippy's palaeoenvironment. If that's not fodder for a blog post, I don't know what is.
Because production time on the mural was short, we decided to augment an existing picture rather than start from scratch. The image in question is below, and was created in 2009 to publicise work by Mike Taylor, Mathew Wedel and Darren Naish on sauropod neck posture (Taylor et al. 2009). The Dorset team liked the image and, though quite dated now, it gave an anatomical and compositional framework that had been approved by several sauropod experts, shaving a lot of time off the production schedule. The final artwork is different to the original in many respects but much of the 2009 DNA remains obvious, including our nod to
Rudolph Zallinger's Age of Reptiles mural.
 |
| PR art for Taylor et al. (2009), showing D. carnegii with its neck held aloft rather than - as was fashionable at the time - held horizontally. 2009 was a long time ago for me, artistically speaking. |
Working with Dippy, and establishing the scene
It's important to any palaeoartwork to know the nature of the actual fossil material behind a reconstruction, and it might come as a surprise to know that 'Dippy the NHM
Diplodocus'* is a different entity to the specimen it's cast from. The 'real' Dippy is Carnegie Museum specimen 84 (CM 84 for short), the holotype of
Diplodocus carnegii, unearthed from Jurassic sediments of Wyoming in 1899. It's a mostly-complete skeleton missing elements of the limbs, the end of the tail and the skull, and these elements were sculpted or casted from other animals to create the mounted Dippy skeletons in museums around the world. This makes Dippy mostly representative of a single individual, but still a composite of several
Diplodocus. CM 84 has been extensively documented - especially in Hatcher's 1901 monograph - and this makes it an excellent specimen to base a palaeoartwork on.
Scott 'Master of Dinosaur Bones' Hartman's 2013 Diplodocus skeletal restoration was used to fill in the proportional gaps, and Tschopp et. al (2015) provided some very useful data on diplodocid osteology, often up close and in clear detail.
*The NHM's CM 84 cast is not the only Diplodocus to bear this nickname: the actual CM 84 specimen was also christened 'Dippy' when discovered in 1899, and several museums around the world use this name for their casts. In this article, my use of 'Dippy' consistently refers to the NHM cast.
 |
| Where the Diplodocus roam: depositional settings of the Morrison Formation at the time when Dippy lived. The Dippy site itself is in southeast Wyoming, among the series of wetlands that line the eastern side of the Morrison basin. From Turner and Peterson (2004). |
CM 84 stems from the centre of the Morrison Formation, a famous Late Jurassic unit that yields, in addition to
Diplodocus, many famous dinosaurs:
Allosaurus,
Stegosaurus,
Brontosaurus,
Ceratosaurus and
Camarasaurus, among others. The Morrison Formation is geographically extensive with major outcrops in Colorado and Wyoming, and additional exposures in 11 other states (above). Palaeoenvironmental studies show variation in climate and habitats across that range. We know that southern regions were drier, that a number of water bodies existed across the basin, and that water and sediment influxes were received from highlands to the west and, possibly, the east (Turner and Peterson 2004). The Wyoming quarry where CM 84 was recovered represents an ancient lake, part of a broader series of wetlands in the east of the Morrison depositional basin (Turner and Peterson 2004; Brezinski and Kollar 2008). It's been suggested that these relatively well-watered settings may have been important habitats for dinosaurs of all kinds, offering abundant plant material compared to the surrounding arid environments (Turner and Peterson 2004). I took these details on board for the mural, changing the backdrop of the 2009 image from a sparse lake margin to a well-vegetated, westward-facing gateway with distant hills. The result is hopefully something not too far off the environment that CM 84 was buried in, and maybe lived in.
Proportions, poses and pedes
Although some tweaks were made to the proportions of the animals from my 2009 image, the basic poses of each was maintained. Readers may question why the necks of the animals have remained aloft when some researchers and artists still use the horizontal neck poses popularised in the late 1990s. The primary basis for horizontal sauropod necks are the famous
Dinomorph digital models (Stevens and Parish 1999 and subsequent works) and, though debates on these matters continue, a number of papers have found issues with these models, to the extent that I'm not sure they're reliable at present. Rather than summarise these issues here, I suggest you simply read Mike Taylor and Matt Wedel's
blog series on sauropod neck posture over at SV:POW! - all the citations and discussion you need are there, and in much greater detail than I could cram into one paragraph.
This means that the postures used in my mural are still, almost 10 years on, based on the conclusions of Taylor et al. (2009). If you missed this paper (which you need miss no more, seeing as it's
open access), it used x-ray data to show that all extant terrestrial amniotes habitually hold their necks with an elevated base during idle but alert behaviour. As summarised by Mike and colleagues in their abstract:
"Unless sauropods behaved differently from all extant amniote groups, they must have habitually held their necks extended and their heads flexed."
In other words, if sauropods didn't carry their necks at an upward angle, they would differ from all terrestrial tetrapods alive today, and there's really no compelling reason to think that was the case. I like this argument because it's based on a broad dataset of real, live animals, not a series of assumptions about how we think they work - when reconstructing animal poses, that's an important distinction. Articulated sauropod fossils show that such poses were attainable, and biomechanical studies suggest that strung-out, horizontal poses would be energetically demanding compared to more vertical poses, and that the necks of sauropods are frankly maladaptive if the neck was not capable of reaching up to gather food (e.g. Taylor et al. 2009; Christian 2010). More work needs to be done here, and it remains difficult to say exactly how sauropods carried their necks for a number of reasons, but data arguing for elevated neck postures seems more compelling than the alternative for the time being. With all this in mind, I am still happy with the neck poses from 2009, and only added some slight curvature to give a sense of motion.
 |
| Of course, no-one is saying that sauropods could only carry their necks aloft: we're talking about their default, habitual pose, not those employed during other behaviours like foraging or drinking. Here's artistic proof. |
But while my sauropod necks remained mostly unchanged, tweaks were made to other anatomies. I missed papers regarding sauropod foot posture in my original work and gave my Diplodocus elephant-like feet, as if they were walking on the tips of their toes. It turns out that this was wrong: their feet were semi-plantigrade and we need to be restoring all sauropods with longer, flatter feet (Bonnan 2005). With sauropod hands having an unusual horseshoe-shaped profile (Paul 1987), it's long past time to bin elephant hands and feet as a model for sauropod appendages: any artists out there still using elephant legs as a model for sauropod limbs, take note. The overall proportions of the animals were modified too, with more muscle added to the neck base, torso and tail base; the cranial proportions corrected, and the torsos given more bulk. I didn't add too much, though: diplodocids were relatively slender as sauropods go, with deep, but not especially wide bodies. They're a world away from the likes of titanosaurs, which were much heftier throughout the trunk (below)
 |
| My PR art for the description of the titanosaur Shingopana songwensis, with another titanosaur - Rukwatitan bisepultus - in the distance. Notice the bulk in their torsos - the chests of diplodocids were a world away from these chunkers. |
A very Dippy face-lift
Ideas about the facial anatomy of sauropods have been undergoing something of a quiet revolution in recent years (as explored in blog posts by
Matt Wedel and
Darren Naish), and good skull material of
Diplodocus allows for artists to consider their craniofacial tissues in detail. Many readers will know that the long-held notion of sauropod nostrils being placed at the top of their skulls has been challenged through careful analysis of their bony nasal anatomy (Witmer 2001). It seems that the obvious nasal openings atop sauropod skulls are only the 'internal' apertures of a larger nasal complex which covered most of their snouts. These are especially obvious in some taxa, like
Giraffatitan (below), but are also evident in diplodocids. Knowing this, we can move the position of the nostrils to the front of the snout, at the anterior limit of the nasal region. This isn't an arbitrary decision: virtually all reptile nostrils are located at the front of their nasal skeleton, so sauropods would be weird if they didn't do this (Witmer 2001).
 |
| Giraffatitan brancai shows us how extensive sauropod nasal skeletons really are - they actually extend right the way down the face (Witmer 2001). Illustration from Witton (in press). |
But other than probable nostril placement, we don't know much about the soft-tissues inhabiting these expanded nasal regions. Were they relatively slender, only slightly modifying the shape of the skull contours, or where they expanded, drastically altering the shape of the face? We don't know, but the unusual noses of monitor lizards give one model for artists to follow. As with sauropods, monitor nasal cavities are large, complex basins occupying much of the snout. Within them sit bulbous cartilaginous nasal capsules, and it's these, rather than the bones of the skull, which create the swollen, sometimes 'boxy' appearance of monitor snouts. If the same was true for sauropods, their facial contours might have deviated markedly from the underlying skull. I used this model in my Dippy mural, adding a healthy bulge of tissue to the face over the entire nasal region. It changes the shape of the craniuim quite considerably, contrasting with the horse-like face so familiar to us in other
Diplodocus restorations, but still - hopefully - being within the realm of scientifically credibility.
 |
| A tiny eye, big nose and Jaggeresque lips. This is not the Diplodocus I grew up with, but all three of these anatomies have a grounding in sound science. |
Regular readers will not be shocked to see covered teeth on my
Diplodocus. The conversation about dinosaur lips and other extra-oral tissues is ongoing, but the presence of covered teeth in virtually all tetrapods suggests we should assume this condition for dinosaurs too, unless we have good reason to remove them (I've blogged on this a lot - see
this,
this and
this). Sauropods meet most of our current, provisional criteria for having covered teeth: their snouts have low foramina counts, which seems to superficially correlate with lips in living species (Morhardt 2009); they lack evidence of sculpting typical of tight facial tissue around their their jaw margins (or anywhere else on their skulls, for that matter - sauropod skulls in general seem to lack obvious epidermal correlates), and their teeth are small enough that they would be easily covered by lips. New data on
Camarasaurus teeth further supports the assertion of generous oral tissues in sauropods (Wiersma and Sander 2017) and, collectively, these lines of evidence suggest a set of (perhaps lizard-like?) lips around the mouth of
Diplodocus is a reasonable inference, without providing any supporting evidence for a perpetual toothy grin.
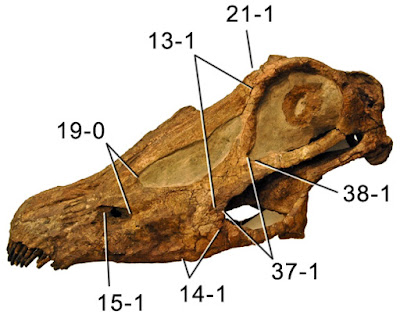 |
| Diplodocus sp. skull CM 11161 - note the well-preserved sclerotic ring in the orbit. It's quite large, but the internal aperture - which the eye peeps though - is pretty small. From Tschopp et al. (2015). |
I was happy to find that we have some good data on eye size in
Diplodocus. Many readers will know that sclerotic rings - small bony plates arranged in a ring that line the front of the eyes of many tetrapods - are great indicators of eye size in fossils. The diameter of the ring itself gives a minimum size for the eyeball, and the internal opening approximates the extent of the visible eye tissue. Tschopp et al. (2015) figure a terrific, only slightly distorted sclerotic ring in a
Diplodocus skull which suggests a reasonably large eye considering the size of the animal, but the ring plates are quite thick, creating a relatively small internal opening. Thus, while the eyeball was large (perhaps indicating good eyesight?), the visible eye area was not huge. No giant eyes for my
Diplodocus in the mural, then, and especially with the additional nasal and oral tissue on the face, they ended up looking quite beady-eyed.
Spines, skin and colour
We don't have any data on the skin for Diplodocus, but skin impressions from other sauropods - including other diplodocids - suggest non-overlapping scales are their most likely covering. I used the extensive skin impressions from the Howe Quarry diplodocid (possibly Kaatedocus?) as my main reference point for the mural: these show not only details of diplodocid scales (polygonal, each about 3 cm across) but also that a line of subconical spines was present along the top of the tail (Czerkas 1992). Some of these were relatively large - up to 18 cm tall - so would be conspicuous even from a distance. These structures were included in the 2009 work and I saw little reason to remove them for the mural, as they remain based on best insight into Diplodocus skin. We don't know how extensive the spine row was in the Howe Quarry animal, so I arbitrarily extended it along almost the entire animal, creating a look consistent the spiny backs of many lizards. The skin was topped of with a number of deep folds: these seem prominent in many living reptiles, but we don't often include them in dinosaur art.
 |
| Colour scheme for my Diplodocus. If you're a carnivorous dinosaur, the body says 'all you can eat', but the tail says 'you can't afford it'. |
Colour and patterning remains a complete unknown for sauropods, so our only mechanism for restoring colour their colour involves looking for modern analogues and considering their likely pigmentation mechanisms. Very generally speaking, larger tetrapods show less striking patterning and duller colouration than smaller ones, and this trend seems common enough to assume it might have been true in fossil tetrapods too (and yes, I know there are plenty of exceptions, but we're looking for the wood here, not the trees). This may reflect, at least in part, the availability of carotenoids - pigments which create bright colours - in terrestrial settings. Animals cannot create carotenoids directly so must ingest them, and the bigger they are, the more they need to generate large patches of brilliant colouration. We know that many birds struggle to attain their maximum degree of pigmentation because terrestrial habitats offer variable, often limited carotenoid availability. If many of these relatively tiny animals struggle to find enough of pigment to colour themselves, it's hard to imagine the biggest terrestrial animals of all time faring any better. If so, sauropods would be reliant on melanin, which animals can synthesise, but only produces dull shades of grey, red, brown and black, and layers of structural colour on their scales. Reptiles employ structural colour frequently to create vivid colours, but mostly in concert with other pigments - green lizards, for instance, have scales with yellow pigment overlain by blue structural colour.
Pigmentation mechanics is not our only consideration, of course: we must also consider colour function. Colour has important roles in animal homeostasis and behaviour, and we have to give our reconstructions colour schemes which are appropriate to their lifestyle and biology. At such large size we might assume that camouflage was not important for Diplodocus, and we might also infer that too much dark pigment would be detrimental to its heat exchange. Dark pigments attract heat, and given that sauropods almost needed to lose heat more than gain it, darker skin have been disadvantageous in hot climates like those of the ancient Morrison. Putting all this together, I chose a fairly dull mottled pattern of browns, creams and greys, with some lighter ornamental scales and spines to break up the monotony. One area that I did elaborate was the tail: if, as long suspected, Diplodocus employed its whip-like tail defensively, it could have drawn attention to its weaponry with colouration and patterning. Eagle-eyed viewers might also note that the smaller Diplodocus has some more vivid patterning, echoing a common condition of reptiles where juveniles are more brilliantly coloured than their parents. I toyed with adding a strikingly coloured juvenile, but decided not to on grounds that tiny, precocial baby sauropods probably didn't hang out with adults, and because less can be more when it comes to composing paintings. Hopefully, the colour scheme is believable and consistent with our understanding of animal colouration, which - 99.9% of the time - is the best we can hope for in the palaeoart game.
 |
| The mural in situ, mere metres from Dippy's tail. This photo was taken at the opening night of the exhibition, hence the funky lighting. The museum is also entirely horizontal, not at a slight angle as shown here, but I'd been at the opening night wine by this point. Note the small panels next to the mural - they explain the science that went into it, effectively being a condensed version of this article. |
That covers the majority of the major decisions that went into the mural, so I'll leave our discussion here. Remember that you have until May 9th to see the mural and other artworks in Dorset. They aren't an 'official' part of Dippy on Tour show so, when Dippy leaves Dorset, the mural and other art won't be following -
book those tickets now if you want to see them. I'll be talking about palaeoart at the museum on March 14th - book tickets for that
here and, if you're reading this and come along, please say hello.
Finally, if you'd like a copy of this mural for yourself, you can grab a good quality print from
my online store, where it's available in a range of sizes. Alternatively, you can access a high quality printable file of the mural if you
sponsor my work at Patreon - details below.
Enjoy monthly insights into palaeoart and fossil animal biology? Support this blog for $1 a month and get free stuff!
This blog is sponsored through
Patreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem a meaningless amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusive Patreon content: regular updates on research papers, books and paintings, including numerous advance previews of two palaeoart-heavy books (one of which is the first ever comprehensive guide to palaeoart processes). Plus, you get free stuff - prints, high quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
- Bonnan, M. F. (2005). Pes anatomy in sauropod dinosaurs: implications for functional morphology, evolution, and phylogeny. In: Tidwell, V. & Carpenter, K (eds) Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana University Press, Bloomington, 346-380.
- Brezinski, D. K., & Kollar, A. D. (2008). Geology of the Carnegie Museum dinosaur quarry site of Diplodocus carnegii, Sheep Creek, Wyoming. Annals of Carnegie Museum, 77(2), 243-252.
- Christian, A. (2010). Some sauropods raised their necks—evidence for high browsing in Euhelopus zdanskyi. Biology Letters, 6(6), 823-825.
- Czerkas, S. A. (1992). Discovery of dermal spines reveals a new look for sauropod dinosaurs. Geology, 20(12), 1068-1070.
- Hatcher, J. B. (1901). Diplodocus (Marsh): its osteology, taxonomy, and probable habits, with a restoration of the skeleton (Vol. 1, No. 1-4). Carnegie institute.
- Morhardt, A. C. (2009). Dinosaur smiles: Do the texture and morphology of the premaxilla, maxilla, and dentary bones of sauropsids provide osteological correlates for inferring extra-oral structures reliably in dinosaurs? Western Illinois University.
- Paul, G. S. 1987. The science and art of restoring the life appearance of dinosaurs and their relatives - a rigorous how-to guide. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present Vol. II. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and London), pp. 4-49.
- Stevens, K. A., & Parrish, J. M. (1999). Neck posture and feeding habits of two Jurassic sauropod dinosaurs. Science, 284(5415), 798-800.
- Taylor, M. P., Wedel, M. J., & Naish, D. (2009). Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica, 54(2), 213-220.
- Tschopp, E., Mateus, O., & Benson, R. B. (2015). A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ, 3, e857.
- Turner, C. E., & Peterson, F. (2004). Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem—a synthesis. Sedimentary Geology, 167(3-4), 309-355.
- Wiersma, K., & Sander, P. M. (2017). The dentition of a well-preserved specimen of Camarasaurus sp.: implications for function, tooth replacement, soft part reconstruction, and food intake. PalZ, 91(1), 145-161.
- Witmer, L. M. (2001). Nostril position in dinosaurs and other vertebrates and its significance for nasal function. Science, 293(5531), 850-853.
- Witton, M. P. In Press. The Palaeoartist's Handbook. Crowood Press.































