|
| Head and neck biting sexual behaviour in Tyrannosaurus rex. A novel, brutal and undeniably speculative reconstruction for tyrannosaurs, sure, but is it the result of pure, unbridled palaeoartistic license, or is there something more to it? |
It seems that "speculation" is the current word on everyone's lips in palaeoartistic circles. Thanks largely to the enormous success of
All Yesterdays, and the recent unveiling of its sequel,
All Your Yesterdays, palaeoartists across the internet have been buzzing with excitement over the possibilities opened by speculative leaps of logic. This is undoubtedly a Good Thing. I wrote
almost a year ago about why I thought
All Yesterdays and the ideas it embodied were great, and a must-see for anyone interested in vertebrate palaeontology or palaeoart. I stand by that, and am certain that many of us in the palaeoart community have be positively influenced by this project in the way we recreate extinct species.
All Yesterdays revelled in speculation about prehistory, arguing that we were not being open-minded enough about our depictions of animal appearance and behaviour. The crux, as anyone reading this probably knows, is that many 'traditional' palaeoart concepts are likely erroneous by being overly conservative, and thus 'fail' at both restoring ancient life and producing convincing looking animals. In addition,
All Yesterdays highlighted a number of conventions which had become tropes within palaeoart, and argued palaeoartists produce far more accurate studies of extinct life when these clichés are broken, not to mention more interesting ones. What gave
All Yesterdays such a strong message was that it, for the most part, was scientifically sound, cleverly turning conventions on their head or showing us logical, plausible ancient phenomena that we'd not imagined before.
For the
All Yesterdays sequel,
All Your Yesterdays, we see a minority of palaeoartists reaching further than it's predecessor dared, showing some very elaborate anatomies and lifestyles which may, in my opinion, go further than reasonable inference, even enhanced with speculation, may allow. Before we get any further, I want to stress that this post is
not a review of
All Your Yesterdays. I enjoyed the book, and think it's well worth seeking out for a look at for some excellent and thought provoking imagery. But yes, it does contain a few images which made me question this newfound speculative approach to palaeoart. We have to bear in mind that
All Your Yesterdays was crowdsourced, the result of a contest for "original, creative concepts that are at least partially in-line with our current understanding of extinct animals" from Irregular Books. This is naturally going to draw a range of knowledge bases, some of which may be more comprehensive than others, and it may be that some of the more eyebrow-raising images therein are simple mistakes. I'm not going to name names here, because I gather the artists behind
All Your Yesterdays were not aware that their work was going to be showcased as a 'significant' addition to the
All Yesterdays canon, but I'll hint that molluscan salinity tolerances, the nesting habits in pterosaurs, the soft-tissues of spinosaurids, hadrosaurs and thyreophorans, and the evolution of viviparity were just some things which prompted this post. It's important to stress that problematic 'overspeculations' are not confined to a few pieces in
All Your Yesterdays, but a small but noticeable chunk of post-
All Yesterdays palaeoartworks which, arguably, jump the palaeoart shark. It's these artworks I want to focus on here.
Getting introspective with speculation
Chiefly, some artwork inspired by
All Yesterdays seems to take license for increased palaeoartistic speculation as a sign that 'anything is possible in nature', without any real consideration for how
likely some possibilities are. Other pieces showcase strange anatomies for the sole purpose of contrasting with more traditional standard depictions, without considering why such reconstructions are common in the first place. These works, presented as part of a movement that I think I understand and agree with, have gone beyond the science which has to underpin any recreation of an extinct being. The question is, how much speculation we can use before our work stops being palaeoart and starts being fantasy images starring extinct species?
 |
| Detail of neck biting Tyrannosaurus. I'm sure he's got a great personality. |
Of course, I'm not the first person to ponder this. Indeed, the inspiration for this post,
All Your Yesterdays,
muses on this same issue:
"In short, speculation in palaeoart should be seen as a sliding scale. At which point does a speculation render itself too extreme? And is it even possible to reach said extreme given the ridiculous soft tissue structures and absurd behaviours present in the modern world? It is, in fact, surprisingly difficult to come up with a speculative piece of palaeoart that is unconditionally ridiculous (at least, so long as the basic rules of anatomy, biology and physics are applied, as they are in science-based reconstructions)."
All Your Yesterdays, p. 7
These words contrast with a few comments online. Amid the near-universal acclaim for
All Yesterdays,
one or
two (and
three,
four,
five) folks have made about palaeoartistic speculation running away with itself, a far cry from the suggestion that palaeoart can never, so long as basic science is followed, be too ridiculous. It seems there's some need, then, for discussion about the appropriateness of speculation in palaeoart: how it should inform our work, how far we can take it, and whether all speculations are equal. After ruminating on this for a couple of days, it seems that the best way to tackle this is by dividing palaeoartistic speculation into three categories (as with any classification of an organic, creative process, are best perhaps viewed as major points a continuum), which I'll call primary, secondary and tertiary. These distinctions effectively denote how far depicted ideas stray from actual data. We'll outline these types of speculation first, and then discuss their use below.
Primary speculations
Speculations directly based on fossil data, whereby the evidence for a behaviour, event or anatomical feature is reasonable, but details may be murky and require some imagination to restore. Gut content, pathological bones and complex track sites are good examples of evidence which can be used to inspire palaeoart using primary speculation. We may not know the entire truth behind these fossils, but we can whittle it down to a few very likely possibilities. Basic elaboration of predicted integument of an animal - making fluffy integuments long or short, altering distribution and so forth - would be an example of primary speculation on anatomy, as would adding things like wattles, skin-folds and other likely anatomical details to reconstructions. With primary speculations, we can be more-or-less entirely confident that we're displaying a degree of truth in our work.
Secondary speculations
Speculations not directly supported by fossil data, but operate within our spectrum of knowledge to maintain a degree of plausibility. This may include extrapolation of common behaviours and, to a limited extent, elaborate anatomies from closely related animals to reconstructed species. Extrapolating some behaviours from or close ecological, anatomical or biomechanical analogues may fall into this camp too. Ritualised behaviours (below), unusual ways of dying and foraging on unexpected food sources are good examples. Depicted behaviours may serve to show the function of prominent anatomies. Slightly unusual interpretations of integument and other body tissues (perhaps as
responses to climate, seasons, sexual selection etc.) probably fall into this category, so long as they are consistent with the integuments known within a 'reasonable' phylogenetic bracket. In short: speculations which adorn fossil species with features so fundamental to animal existence that, even in the absence of fossil data, we can be confident they occurred in deep time.
 |
| Ritualised courting chaoyangopterid pterosaurs, Lacusovagus magnificens. Did pterosaurs do this? There's no direct fossil evidence for it, but the abundance of ritualised mating behaviour in modern animals suggests we can be relatively confident that they, and other fossil species, used complex ritualised behaviour. This undoubted speculation gains indirect support from the broad array of sociosexual devices we see on many fossil species, and hints of ancient sexual dimorphism, both of which indicate sexual behaviours were as complex and sophisticated in prehistory as they are today. Image from Witton (2013). |
Tertiary speculations
Speculations operating completely outside, and sometimes contradicting, fossil data. May rely entirely on application of very specific modern animal behaviours and anatomies to fossil species, often transferring rare, sometimes highly specialised lifestyles to fossil animals. There is no particular logic or reason behind these applications: they are entirely arbitrary. In other cases, complex biologies and life histories are invented for fossil taxa. Creation of soft-tissue anatomies without, or in spite of, consideration of underlying musculoskeletal system and/or soft-tissue fossil data. Reliant on the absence of data concerning fossil species, because 'anything is possible'. Hypothetical examples of such speculations are things like lactating dinosaurs, notosuchians with trunks, an egg-laying
Deinotherium, hadrosaurs with antler-like structures growing atop their crests. Jaime Headden's
woolly ankylosaur, his cautionary
'mess of speculation', is a knowing graphic example of tertiary speculations gone mad.
Speculations, what are they good for?
If these are the tools of the speculative palaeoartist, what are their application? Anyone familiar with palaeoartistic practises will recognise that the former two grades of speculation are standard tenets of palaeoart. Such speculations provide our leaps of logic into prehistory and, without them, palaeoart would be an pretty limited endeavour, probably entirely formed of musculoskeletal reconstructions. It's important to recognise that such speculations were not originated by
All Yesterdays, as primary and secondary speculations have
always been used in palaeoart. The masterstroke of
All Yesterdays was to show how primary and secondary speculations could be bolder and more imaginative than most mainstream palaeoart suggested. The result is artwork which is both interesting, unique
and supported by actual data.
The image at the top of this post is the result of such an inference. It's well known that many large theropods engaged in head-biting behaviour, and some specimens of
Tyrannosaurus (including BHI 3033, better known as the common
T. rex museum mount 'Stan') bear particularly extensive damage to their posterior skulls. The inference made here is that
Tyrannosaurus engaged in aggressive head and neck biting during copulation, a widely seen behaviour among vertebrates that can often involve substantial damage to the head and neck of the female, sometimes leading to death. I'm not the first to envision this behaviour for tyrannosaurids. Tanke and Currie (1998) suggested nuptial biting as a cause of tyrannosaurid head pathologies but suggested it was refuted by the apparent small size (50% of full size) of many tyrannosaurids with head wounds. Of course, it now seems that dinosaurs became sexually mature when only half grown (Erickson et al. 2007), so this hypothesis may be back on the table. The resultant image is a radical and speculative depiction of
Tyrannosaurus behaviour, but one that has a foot firmly set in science.
 |
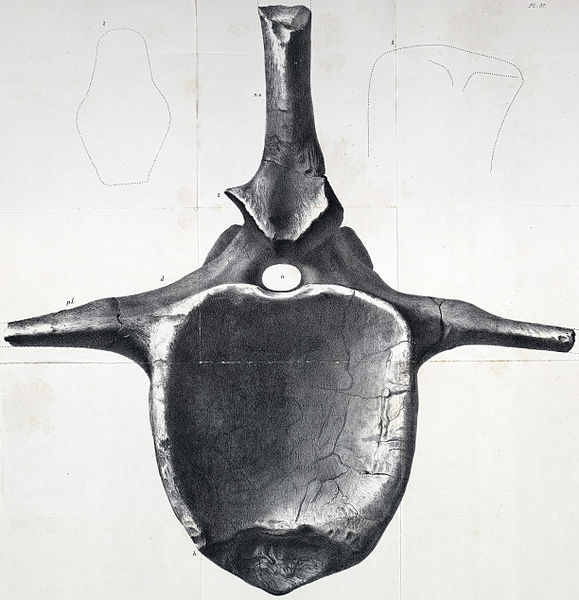
| Cast of the skull of Tyrannosaurus 'Stan', BHI 3033, at in the Oxford University Museum. Stan's skeleton is particularly damaged around the posterior head and neck region, with a probable tooth wound penetrating it's braincase, a smashed postorbital bar (a dorsal projection of tyrannosaur skulls which anchored neck muscles) and broken neck vertebrae. Photograph by Marc Vincent, from Love in the Time of Chasmosaurs. |
The same cannot be said for tertiary speculations. Some inferences made at this level are so far removed from actual data that they have little or no evidence to support them, and thus abandon the scientific basis which should underpin any palaeoart. Others may disagree, but I think good palaeoart, like good science, is led primarily by evidence, not speculation. This most obviously impacts tertiary speculations which arise, it seems, for the sole purpose of overturning convention. "This animal is always shown like/doing this... what if it looked like/did THIS SHOCKING THING?!??" While there's nothing wrong with trying to keep palaeoart fresh, we should remind ourselves that not everything common in palaeoart is a
trope or meme, or the product of unimaginative artists. Sometimes, that's just how animals were, and conventions are based in very sound evidence. Deviating from these conventions is a move away from data, which is the exact opposite of what we want to be doing here.
Other tertiary speculations apply highly unusual behaviours borrowed from modern animals, or those which are entirely made up, to fossil species for no clear reason. This can be effective on occasion, presenting a fossil species in a radical light which may make us reconsider our preconceived notions of that species, but I'm generally not a fan. Why, of all the behaviours that we can imagine or observe in in the modern day, should we chose that specific animal as a model? And do we really expect the rarest, most elaborate and weirdest behaviours to be present in specific fossil animals? Are we actually predicting that extinct animals behaved (often adorned with the same colour schemes and patterns) exactly like these aberrant modern animals? We'll score far more science points if we apply more widespread behavioural phenomena to our palaeoart. This doesn't mean we have to confine ourselves to dull behaviours like travelling and foraging, because we can also rely on primary and secondary-level speculations to give us behaviours like resting, taking care of personal hygiene, reproducing, interacting with one another and so on. Likewise, lots of interesting anatomies can be extrapolated from the fossil record itself. In sum, while we should take inspiration from modern taxa, arbitrarily 'transforming' fossil animals into ancient versions of modern species stretches credibility quite far, and is perhaps a rather unscientific approach to our work (this point echoes one made earlier, also in response to some art in
All Your Yesterdays, at
Laelaps).
A counterargument could be made that tertiary speculations allow us to imagine how sophisticated and complex ancient worlds were but, again, I question this. Like any guesswork, they're of questionable significance. Unknowns are unknowns. Tertiary-grade restorations are as likely to be incorrect as accurate. These depictions may fire the imagination briefly, but the flames are tiny compared to those fuelled by cool ancient behaviour derived from actual evidence. It's one thing to see a shocking piece of palaeoart, but quite another to realise that there's actually tangible evidence behind it. Rather than pondering the great unknowns of deep time when confronted with a tertiary speculation, I frequently react with the opposite approach, thinking about what we can actually deduce about a given issue, and what a more likely interpretation may be.
 |
| Why I find tertiary speculations frustrating. The fossil record is full of interesting animals with known interesting behaviours, like these burrowing Oryctodromeus, and yet they are frequently overlooked in palaeoart for entirely speculative renditions of familiar taxa. Check out this post for more on this animal and it's need for a PR campaign. |
This brings us to a more pragmatic bugbear about tertiary speculations. Extremely speculative palaeoartworks are actually fairly common, at least online, while innumerable cool palaeontological topics with a significant factual basis are completely ignored. Why use art to make rather hollow points about unknown topics when there's plenty of art to be made concerning subjects we
do know about? Even familiar animals have unusual, rarely-depicted behaviours which we can infer from fossils with minimal amounts of speculation (such as the tyrannosaurs above), not to mention the shedloads of fossil species which are wholly unrepresented in art (and yes, I'm well aware of the hypocrisy of saying this in a post featuring
Tyrannosaurus), many of which are also known to have interesting and unusual behaviours. Heck, it's common knowledge that palaeoart is heavily biased towards a few taxa, so just showing some of these rarely seen animals would be a thought-provoking, cliché-busting achievement in itself. Is it not better, as scientific illustrators, to base our work on what we know rather than what we don't?
Which leads to...
So, yes, despite being an advocate of using speculation in palaeoart, I'm not a huge fan of the
extreme and
uncontrolled speculation we're seeing creeping into modern portfolios. This may sound like I'm jumping off the
All Yesterdays bandwagon, but I don't think I am. Most of our best palaeoartists - including those behind
All Yesterdays - use speculation of primary or secondary grade, and are more notable for avoiding clichés and artistic conventions than they are for presenting highly speculative lifestyles and anatomies in fossil species. They elaborate existing knowledge to create more convincing depictions of fossil animals, and apply detailed research of the fossil record to show us sights we've never seen before. Some of their work may seem outlandish and brash, but it's actually far more measured than it looks.
I'm sum my point up as this. While we should be using speculation to push palaeoart to its limits, we need to know both which bits we can push, and when to stop before our speculations get the better of our work. This doesn't deny us licence to make our reconstructed ancient worlds amazing and interesting and, in fact, it may make our work more striking. It's one thing to see an outlandish reconstruction of the past, but all the more poignant when we realise the weird, strange or even shocking visage before us is based on truths, and not just imagination.
References
- Erickson, G. M., Rogers, K. C., Varricchio, D. J., Norell, M. A., & Xu, X. (2007). Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition. Biology Letters, 3(5), 558-561.
- Tanke, D. H., & Currie, P. J. (1998). Head-biting behavior in theropod dinosaurs: paleopathological evidence. Gaia, 15, 167-184.
- Witton, M. P. (2013). Pterosaurs: Natural History, Evolution, Anatomy. Princeton University Press.







.jpg)