
 |
| Quetzalcoatlus lawsoni scavenges a juvenile Torosaurus, recently killed (judging from the decapitation) by a large tyrannosaur. We've been waiting for ages to learn more about this pterosaur following a near-50 year delay in the description of its remains, but we're finally there! So what have we learned about Quetzalcoatlus proportions and anatomy from this glut of new information? Read on... |
Almost a full half-century after its discovery, December 2021 finally saw the publication of a suite of technical papers on one of the most famous of all pterosaurs: Quetzalcoatlus. An entire themed collection of Quetzalcoatlus articles have been bound together in a special Journal of Vertebrate Palaeontology memoir, and they’re all — happy days — open access. This is great news for everyone with an interest in this animal or pterosaurs in general, and it represents a much-needed turnaround of a long-standing embargo on the Quetzalcoatlus material. Several authors have been attached and let go from The Big Quetzalcoatlus Project since the early 1970s and those of us without access to its fossils have done what we can to understand it from snippets of information published here and there. The memoir team — who tackle Quetzalcoatlus anatomy, systematics, palaeoenvironment and functional morphology — have thus ended a frustrating half-century-long wait for more information on this hugely popular, much-loved pterosaur. I have no doubt that these new papers — particularly Brian Andres and Wann Langston Jr.’s super-detailed and extensive osteology (Andres and Langston 2021) — will turn Quetzalcoatlus into a modern cornerstone of pterosaur research. Essentially overnight, the memoir team have turned Quetzalcoatlus from an almost non-entity in the scientific literature to an embarrassment of pterosaurian riches, and I look forward to the many new insights their work will foster.
There’s much to say about the entire volume and if you’d like a full run-down you should check out Darren Naish’s overview at Tetrapod Zoology. Here, I want to focus on one specific paper: the overview of functional morphology written by Kevin Padian et al. (2021). Many readers will have seen details on this paper in the media because it’s the one that turns our piles of Quetzalcoatlus fossils into a living, breathing animal, opining on what it looked like and how it walked, flew, and foraged. In other words, the kind of stuff that many people want to know most about these awesome animals. It's the paper that's drawn most of my attention because of my own research interests in azhdarchids: chiefly their palaeoecology, functional morphology and reconstruction (see Witton and Naish 2008, 2013; Witton and Habib 2010; Naish and Witton 2017 for examples).
And to cut straight to the chase, I want to talk about this because I suspect Padian et al.'s paper is destined to be the most controversial of the memoir’s contributions. It contains a lot of ideas and opinions that will be classed as unusual, non-mainstream takes on pterosaur palaeobiology and while some are novel, others are resurrected from papers written by the senior author decades ago (e.g. Padian 1983a, b, 1988, 2003, 2008). I say 'resurrected' because some of the ideas in question have since been rebutted or struggled to gain wider acceptance among pterosaur workers, such that reading Padian et al. (2021) has a distinctly vintage feel, like it's been beamed in from 20 or 30 years ago. I'm specifically interested in their handling of Quetzalcoatlus proportions, ground posture, wing folding and flight mechanics because Padian et al. make some genuinely strange suggestions around these topics; Quetzalcoatlus is restricted to crouching poses, might have flown with its legs tucked under its body, had its principal wing membrane attached to its hip, and potentially took to the air with a bipedal leap.
Proof that Padian et al. is destined to be controversial is found in the paper itself, where we find in-text admissions that the authorship team could not agree on matters of launch mechanic, flight posture and wing membrane configuration (in all instances, palaeontologist Kevin Padian is stated as disagreeing with his coauthors, biomechanicist Jim Cunningham and palaeoartist John “All Yesterdays” Conway). On top of this, some presented data conflict with other parts of the memoir, and there are a few potential errors that affect the reliability of the paper’s conclusions. It is, in detail, something of a wild read, and reaction to this paper is going to be mixed among pterosaur experts. Nevertheless, Padian et al. (2021) will also be a potential source for pterosaur reconstructions for the foreseeable future, and given that both the paper and associated press coverage are publicly accessible, I think it's right to have some responses from pterosaur workers online, too. In that vein, I want to point out which ideas should be considered unusual, which might jar with the rest of pterosaur research, and highlight a few issues I’ve identified when combing the paper. I hope the following is taken in the spirit it’s intended — an honest response to a paper on an animal I’m deeply interested in — especially because two of the authors (Jim and John) are good friends. John is also aware of some of my misgivings so the following won’t come as a complete shock. In any case, I hope it’s clear that the intention here is not to whale on new research, but to highlight areas I predict will be contentious or did not find compelling, on the chance that they are of interest to others who find Quetzalcoatlus fascinating.
Old vs. new: Quetzalcoatlus 2021 compared to older reconstructions
 |
| My 2016 skeletal reconstruction of Q. lawsoni (then known as 'Q. sp') is now consigned to history. How did this now-six-year-old reconstruction hold up to the might of The Memoir? Read on... |
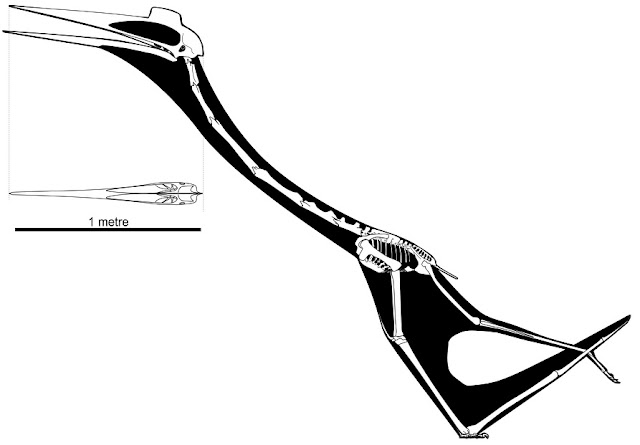
One of the things I was keenest to see in Padian et al. (2021) was how my various reconstructions of Quetzalcoatlus have fared against new data. I’ve designed a lot of azhdarchids, including Quetzalcoatlus, for film and TV and wanted to see how close I’d landed to the reality of this animal using scraps of information gleaned from other papers. The artwork in Padian et al. (2021) is both copious and all excellent, stemming — of course — from the hand of John Conway, and we’re treated to a lot of skeletal reconstructions in multiple views. The paper largely focuses on Q. lawsoni, the smaller of the two named Quetzalcoatlus species, because virtually the entire skeleton of this animal is represented across multiple, similarly-sized specimens. As we've known since the 1970s, the charismatic giant wing that represents the giant Q. northropi can only take you so far in understanding Quetzalcoatlus: Q. lawsoni is really where the action is. And one thing to point out straight away is that our Q. lawsoni material is from several similarly-sized animals, but that there’s a lot of variation in limb metrics across them. They all seem to reach 4.2-ish m wingspans through similar, but slightly different proportions (Andres and Langston 2021). This is interesting for all sorts of reasons, but also complicates any attempt at reconstruction. Probably the most obvious solution is to figure out what an ‘average’ Q. lawsoni looked like and work from that, although it would be neat to compare the extremes of proportion across the dataset too (something we're not doing here today).
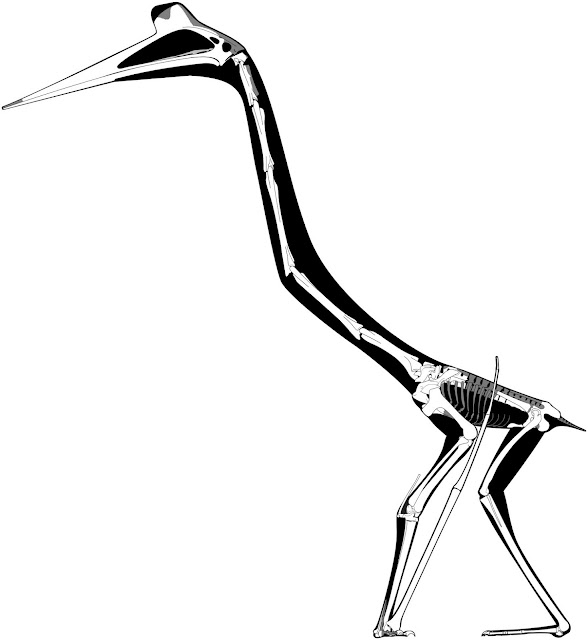 |
| The Padian et al. (2021) reconstruction of Q. lawsoni "posed in quadrupedal terrestrial stance". The half-crouched limbs are not artistic whimsy, but tied into ideas of Q. lawsoni hindlimb motion. We're going to get into that in a moment. |
Comparing my old reconstructions with the new data, I think (if I may say so) that my work stands up relatively well. In honesty, I was surprised by Padian et al.’s (2021) assertion that: “there has never been a justification for the proportions of the bones used in any [Quetzalcoatlus] illustration”. This simply isn’t true because, despite the embargo over Quetzalcoatlus material, a lot of information on Q. lawsoni has been published over the last 50 years. These include mostly accurate limb metrics (Unwin et al. 2000); a full skull description (Kellner and Langston 1996) and dimensions of the cervical vertebrae (Steel et al. 2007; Witton and Naish 2008). There have even been pretty decent skeletal reconstructions based on examination of the original fossils (Paul 2002). These data are why the skeletal reconstruction published by Padian et al. (2021) isn’t massively different to some carefully researched pre-2021 versions. Compared to my own work, the only major discrepancies I found concern some posterior cervical lengths, the length of the body, and the size of the wing metacarpal. On the latter, my skeletal used a 620 mm length derived from Unwin et at al. (2000), which it now appears is too long: actual Q. lawsoni WMC lengths were in the range of 420-470 mm. Overall, Quetzalcoatlus was a little shorter in the arm than I’m used to, and fractionally longer in the neck and body, but it’s not a total visual transformation. Other distinctions between my older work and John’s new skeletal are just matters of opinion. For instance, the Padian et al. Quetzalcoatlus has a very tapered posterior skull, which I think is unlikely given the general condition of azhdarchoid crania. Specifically, completely known skulls from the azhdarchid Zhejiangopterus linhaiensis and at least one member of the azhdarchid sister clade, Chaoyangopteridae, have tall, sheet-like frontoparietal bones extending over and beyond their braincases (Cai and Wei 1994; Lü et al. 2008), and we see similar conditions in thalassodromids/ines as well. I thus regard this condition as likely for Q. lawsoni, but this will remain nothing more than opinion until we find a more complete skull.
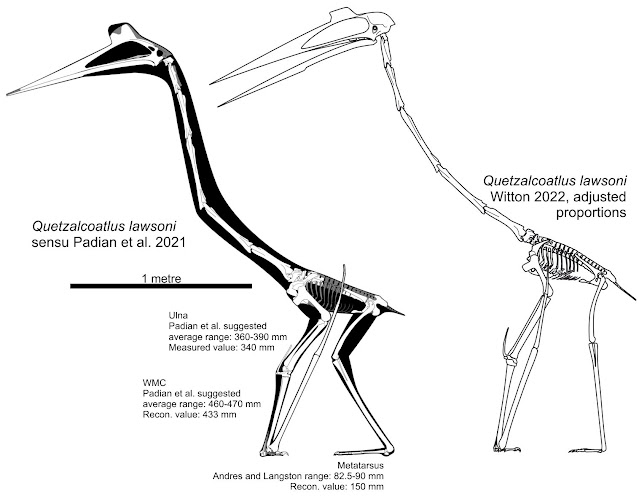 |
| Is the crouching pose of the Padian et al. Q. lawsoni reconstruction necessary? Ignoring the hindlimb restoration philosophy (see below), there may be scaling issues with the forelimb affecting things too: an 'adjusted' skeletal to the right shows that Q. lawsoni could stand tall without issue. |
But in checking out the new reconstructions I also noted some less subjective differences. One of the more striking aspects of Quetzalcoatlus 2021 is the proposed habitual crouching pose. It reflects both assumptions about the hindlimb articulations (which we’ll discuss at length below) as well also the unexpected shortness of the forelimb. But even accounting for that short wing metacarpal, the wing looked strangely stunted to me. Upon investigation, I found that the wing skeleton is probably incorrectly scaled. Specifically, when compared to metrics given in Padian et al. (2021) and Andres and Langston (2021), the reconstructed radius/ulna and wing metacarpal lengths are 11 and 10% shorter (respectively) than an ‘average’ Q. lawsoni wing. This makes the arm quite a lot shorter than it should be and, when adjusted, there’s no problem making Quetzalcoatlus stand in a more typical, fully-upright posture. Indeed, the forelimb becomes long enough that the entire hindlimb can be extended vertically under the body without the arm looking over-extended (above).
The size of the foot also drew my attention. It’s been remarked that azhdarchids had small feet (Cai and Wei 1994; Hwang et al. 2002; Witton and Naish 2008; Andres and Langston 2021) and yet Padian et al. (2021) show Quetzalcoatlus as a relatively large-footed animal. The diminutive foot size of azhdarchids was one reason Darren Naish and I suggested they were terrestrial foragers back in 2008 (Witton and Naish 2008) and when I saw the big, flappy feet of 2021's Q. lawsoni I thought we’d got things wrong. But, again, there’s a measuring complication here. Padian et al. (2021) suggest the metatarsus (the shaft bones of the foot) was about 150 mm long, which is about 25% of the tibiotarsus length, and this is what's shown in the reconstruction. But Andres and Langston (2021) record the metatarsus as only 15% of the tibiotarsus, and 82.5-90 mm long. Andres and Langston further stress that Quetzalcoatlus had the third-shortest foot, relative to body size, of any known pterosaur, and this emphasis makes me think their measurements are more likely to be correct. If so, and we then assume — as suggested by Padian et al. — that the toes were a similar length to the metatarsals, Q. lawsoni would have had tiny feet of c. 160-180 mm long. This is a little over half of what's reconstructed for the Padian et al. restoration, but similar to the foot proportions of Zhejiangopterus.
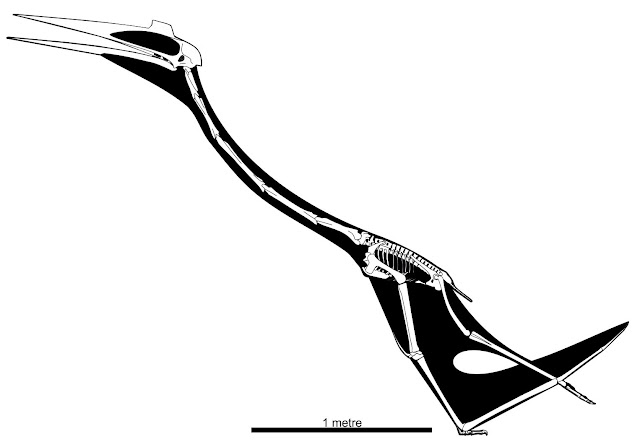 |
| My 2022 skeletal reconstruction of Quetzalcoatlus lawsoni, incorporating the adjusted proportions outlined above. |
Once Quetzalcoatlus 2021 is adjusted to suit these adjusted measurements, it looks a lot less strange. You can get a sense of this from my own rebuilt skeletal reconstruction of Q. lawsoni, above. It should be stressed that there is a defence to these scaling issues: the aforementioned variation in limb metrics where specimen proportions can vary by over 10%. So perhaps Padian et al. haven’t reconstructed an ‘average’ Q. lawsoni, but they’ve still reconstructed something within the proportions of this species? There may be some validity to this, but comparing the presented reconstruction to the metrics of Andres and Langston (2021) suggests it's still something of a stretch. Nevertheless, the weird variation in Q. lawsoni proportions may be where all these issues originated.
Deja Qu, part I: crouching Quetz, hidden controversy
Let’s now look beyond proportions to functional morphology, starting with that strange crouching hindlimb. This reflects the idea that the Quetzalcoatlus femur was perpetually held subhorizontally with a maximum downward rotation of only 70-75𝆩 (Padian et al. 2021). This, it's said, prohibits the femur from swinging backwards under the pelvis as is widely interpreted and illustrated for pterosaurs across scientific literature and palaeoartworks, and it's not a new idea: it's taken straight from Padian papers published in the 1980s. The arguments are principally the same, too: that the articular surfaces of the pterosaur knee do not allow the leg to straighten, and that the femoral curvature of pterosaurs recalls that of birds, implying a subhorizontal orientation (see Padian 1983a, b; Padian et al. 2021). Padian et al. (2021) also mention that their proposed posture scores points for fitting Jurassic pterosaur tracks from Crayssac, France, an idea that further ties into classic Padian literature. Why fit Quetzalcoatlus into the tracks of relatively tiny Jurassic pterodactyloids and not the Haenamichnus tracks widely considered to have been made by a Korean azhdarchid (Hwang et al. 2002)? Because Kevin Padian (2003, 2008; Padian and Olsen 1984) has long been sceptical about the origins of pterosaur tracks, cumulating in the belief that only examples found in southern France are genuine pterosaur ichnites. Most or all others, he argues, were left by other reptiles; chiefly, crocodylian-like ones.
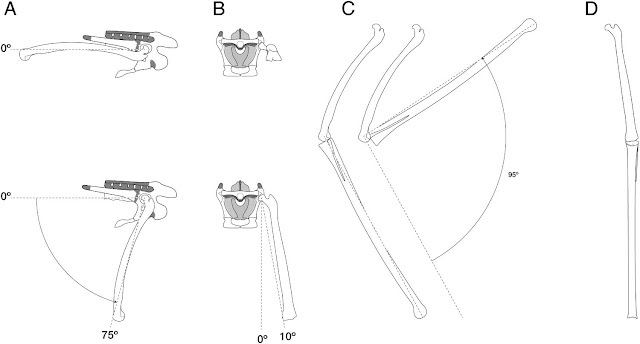 |
| Suggested ranges of motion at the Q. lawsoni hip and knee, according to Padian et al. (2021). The precision drawings and figures suggest a lot of confidence in these data, but they contrast with several comments about the poor quality of the Q. lawsoni pelvis in the memoir, and the difficulty of reconstructing it accurately. The proposed range of knee articulation is also very restricted compared to analyses of this joint in other pterosaurs. |
Understanding that Padian et al. (2021) has been written from this perspective explains why its discussion of hindlimb mechanics frequently jars against more recent studies. Padian et al. concede that the concept of subhorizontal pterosaurian femora contrasts with the conclusions of at least one team (Costa et al. 2014), but don't mention the heaps of other investigations it also conflicts with (e.g. Bennett 1990, 1997, 2001; Unwin 1996; Fastnacht 2005; Wilkinson 2008). While it would be incorrect to say that we understand everything about the motion of the pterosaur hindlimb, most researchers are pretty happy that the femur could swing into a subvertical pose. Indeed, some studies conclude that this is the optimal position for the pterosaur hindlimb when walking, providing the best mechanical advantage for the muscles that move the leg forward and back (Fastnacht 2005; Costa et al. 2014). It’s also generally observed that the articular surfaces of pterosaur knees extend to the tips of the limb bones, allowing them to adopt almost entirely straightened knee poses (e.g. Bennett 2001; Wilkinson 2008). Padian et al. (2021) provide the first assessment of this for azhdarchid knees, but I admit to wondering why they think the condyles are so limiting when azhdarchid hindlimb joints look pretty similar to those of other pterodactyloids (see Godfrey and Currie 2005; Averianov 2010, and Andres and Langson 2021 for images).
With the weight of opinion being that pterosaur hindlimbs were actually pretty different to bird legs, I'm surprised the Q. lawsoni functional analysis leads so strongly with its assessment of a bird-like subhorizontal femur. At very least, those other studies warrant discussion. And as for the seeming validation that a crouching Quetzalcoatlus can be made to fit Jurassic pterosaur trackways, this is a moot point: conventional, upright hindlimb postures fit these tracks too (Bennett 1997; Mazin et al. 2003). The take-home here is that the proposal of Quetzalcoatlus having a subhorizontal femur, and thus being limited to a strange, crouching pose, is both odd and not well substantiated against the consensus view of pterosaur research. It really needs bolstering with more data to be credible.
Deja Qu, part II: Leg folding…
I mentioned above that some disagreement exists among the Padian et al. (2021) team on several topics, two of which concern flight pose and membrane shape. While Jim and John advocate something approximating the classic sprawled-leg flight pose and at least some degree of hindlimb membrane attachment, Padian prefers a bird-like configuration where the hindlimb is tucked underneath the body and the wing membrane anchors at the hip. Again, these latter ideas are Padian hypotheses that first aired 40 years ago (e.g. Padian 1983b; 1988). Predicting pterosaur membrane shapes remains a complex issue and is beyond our scope for discussion here: it’ll suffice to say that there is no evidence for a pelvic membrane attachment in any pterosaur, and that the handful of inboard membrane fossils we have collectively point to a distal hindlimb attachment across Pterosauria, including in Azhdarchoidea. This was well documented by Ross Elgin et al. (2011), a paper which Padian et al. cite and (probably unfairly) dismiss with just a few words. As for the question of flight pose: this boils down to whether Quetzalcoatlus was incapable of adopting the classic ‘sprawled-leg’ posture widely reconstructed for pterosaurs and, if not, did it have to adopt an unusual, avian-like one instead?
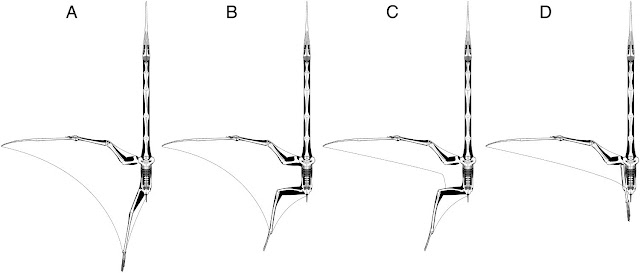 |
| Q. lawsoni wing poses illustrated by Padian et al. (2021): which do you prefer? John Conway and Jim Cunningham are on record preferring model C, while Kevin Padian argues for D. My vote, given what fossils show of pterosaur wing membranes, would be for something between B and C (distal hindlimb membrane anchor, but a tighter trailing curve than B). |
As alluded to above, discussions over pterosaur pelvis-hindlimb arthrology are nothing new. How far pterosaurs could move their femora around has been the subject of a large number of papers (e.g. Padian 1983a, b, 2003; Wellnhofer 1988; Bennett 1990, 1997, 2001; Unwin 1996; Wilkinson 2008; Costa et al. 2014; Frigot 2018) leading to a general consensus that most or all pterosaurs could move their hindlimb through a wide range of motion, walking and standing with a near-vertical femur but also swinging their legs out in flight. This conclusion is not just based on manually articulating bones but also on hundreds of articulated fossils showing pterosaurs preserved with both upright and splayed hindlimbs. These include at least two Zhejiangopterus specimens with butterflied hindlimbs (illustrated in Cai and Wei 1994 and Witton 2013) that show azhdarchids conforming to pterosaur norms. If Quetzalcoatlus was incapable of adopting a hindlimbs-out flight pose, it would have been highly aberrant and we’d need good evidence of such an interpretation: ideally, a well-preserved pelvis with an uncrushed acetabulum (hip socket) and a correspondingly well-preserved femur that allowed us to demonstrate, beyond doubt, limited capacity for hindlimb abduction.
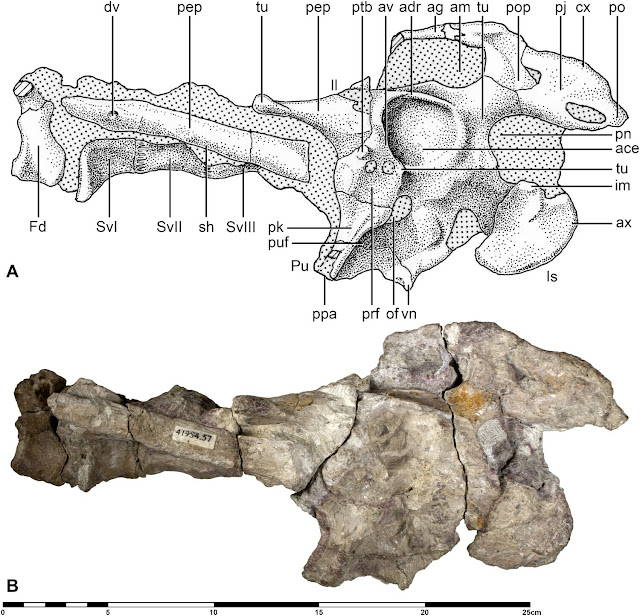 |
| The Q. lawsoni pelvis as illustrated by Andres and Langston (2021): it's far from the best-preserved piece of our Q. lawsoni inventory. Are these the sort of pelvic remains we can use to substantiate a radical departure from our typical interpretations of pterosaur hindlimb arthrology? Probably not. |
But, alas, here’s how Andres and Langston (2021) describe the only recovered pelvic material of Q. lawsoni: “fractured, heavily encrusted with concretionary material, and [with] matrix… often stained a similar color to the bone.” They conclude that “this pelvic plate is not preserved well enough to decisively determine its orientation with respect to the vertebral column” and that the angle of the acetabulum cannot be interpreted with certainty. Even Padian et al. (2021) concede that “the pelvis cannot be reconstructed in three dimensions with confidence”. There's an agreement, then, that we can't reconstruct the three-dimensionality of the Q. lawsoni pelvis without doubt, and this is a problem. We've learned from multiple studies that restoring pterosaur leg mobility is influenced by numerous factors including the shape and orientation of the hip socket, the precise angle of the pelvis with respect to the spinal column, the articulation of the pelvic bones themselves, and the inclination of the torso (e.g. Wellnhofer 1988; Bennett 1990; Fastnacht 2005; Wilkinson 2008; Costa et al. 2014). In other words, we need really, really excellent fossils to even start thinking about such investigations and if we can't reconstruct the Q. lawsoni pelvis, we cannot say much about the range of motion of the leg.
It's for this reason that Padian et al. can only infer avian-like hindlimb mobility for Q. lawsoni, which they openly declare in their introduction to this topic: “Given the bird-like features of the entire hind limb, which not only bear anatomical resemblance but speak to functional similarity, it appears reasonable to begin with the kinds of postures and degrees of movements found in birds”. This is not the right approach and certainly undermines their abstract assertion that "In flight, it is most plausible that the hind limbs were drawn up bird-like, with the knee anterior to the acetabulum". Surely, if we can't model the hindlimb arthrology for Quetzalcoatlus, we have to fall back on what we're learned from other pterosaur species, not point to an anatomically distinct, phylogenetically distant pterosaur relative and made sweeping inferences? In all, I find nothing compelling about the concept of Quetzalcoatlus having to tuck its legs up like a bird, and I 100% agree with Jim and John in their endorsement of a more traditional, hindlimb-splayed flight configuration.
…and wing folding
Discussing flight brings us to another potentially contentious topic: the Q. lawsoni wing, or, rather, wing folding. We know a fair bit about how pterosaurs collapsed their wings for standing and walking (e.g. Wellnhofer 1988; Unwin 1996; Bennett 1997, 2001; Wilkinson 2008) and the general conclusion is that pterosaur forelimb articulation was complex. Their arm joints didn’t articulate uniaxially (i.e. in one plane); instead, the elbow and wrist deflected their distal limb bones medially and laterally as they opened and closed. We've found that, to get pterosaurs walking in their trackways, the pterosaur elbow needed to stick out from the body a little and that (like many dinosaurs) the palms of pterodactyloid hands faced inwards, as if the hands were ready to clap for a round of applause*. This is why pterodactyloid trackways show handprints with sideways projecting fingers: the digits have swung under the big knuckle of the wing digit to extend away from the body. The wing finger itself follows the same rules, so it folds up along the outside of the wing. A quirk of the wing metacarpal joint means that the wing digit is somewhat posteriorly deflected when it does this, stowing alongside the forearm during terrestrial progression. These basic findings are something that we’ve modelled from pterodactyloid bones and also witnessed in dozens, maybe hundreds, of well-preserved pterosaur fossils. Whenever we have an articulated, tightly folded pterosaur wing, the wing finger lies over the radius and ulna, not under it, and the palm of the hand faces inwards. We know this applies to azhdarchids too, thanks to articulated fossils of Zhejiangopterus (Cai and Wei 1994). We also have azhdarchid tracks, Haenamichnus, showing their hands were orientated in a typical, ‘palms inward’ pterosaur fashion when walking (Hwang et al. 2002). All expectations are, therefore, that Quetzalcoatlus would follow this familiar configuration.
*The situation is different in non-pterodactyloids, but that’s another story.
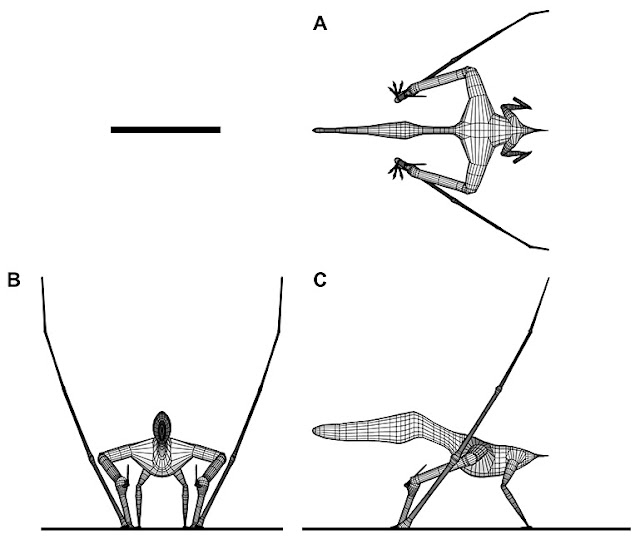 |
| One of my favourite images for showing the complexity of pterosaur forelimb articulation, from Wilkinson (2008). The pose here can be regarded as 'extreme' as we have good data indicating that pterosaurs stood more upright than this, but the orthographic views show how the forearm and hand are deflected as the wing folds. Note how the wing finger and walking fingers rotate around an axis parallel to the midline of the body, allowing the fingers to project sideways while the wing finger folds against the outer arm. |
Given these relatively well-established models, it’s something of a surprise to see Padian et al. (2021) showing Q. lawsoni doing something different. The wing is positioned so that the palm of the hand faces somewhat forward, allowing the wing digit to tuck under the elbow, despite the walking fingers still projecting laterally. The ability to draw the wing finger under the elbow is, apparently, a consequence of a slight downward deflection to the end of the wing metacarpal which changes the orientation of the joint, but I suspect it was also influenced by the methodology for modelling the standing pose. It’s reported that this was deduced by manually positioning casts of Q. lawsoni fossils in a plausible upright arrangement, an exercise which "began by placing the distal end of the wing metacarpal... with its distal condyles oriented posteriorly (so that the wing finger could be directed behind the elbow and close to the body wall)". It seems it was decided, a priori, that this is where the wing finger should go.
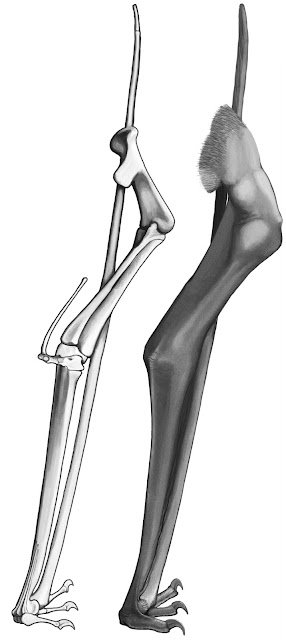 |
| Proposed wing folding of Q. lawsoni, from Padian et al. (2021). |
It’s a shame that no photos or diagrams of this work were published because, while those of us who have not handled the Quetzalcoatlus bones can’t really say that this interpretation is wrong, there are lots of legitimate questions about it that make me hesitant in accepting it outright. For example, Quetzalcoatlus is not unique for having that slight downturn at the end of the wing metacarpal: we see similar conditions in taxa like Pteranodon and Tapejara. In Pteranodon at least, they’ve been factored into arrangements of the folded pterosaur forelimb and do not result in the wing finger tucking under the elbow (Bennett 2001). I'm also not clear how the Q. lawsoni walking fingers are depicted as splaying out to the side when their respective metacarpals are positioned on the front of the wing: unless these joints were strangely bevelled, surely they should be facing more posteriorly? It's also strange to have the palm facing forward at all, as other studies exploring the impact of angling the pterosaur palm forward find that such poses are only possible if the forelimb adopted a crazy, implausible configuration (Bennett 2001).
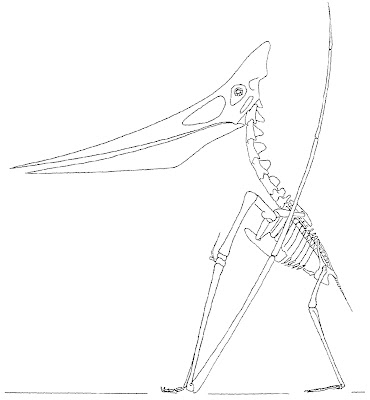 |
| Exploring how pterosaurs stood has taken us down some strange roads. In 2001, Chris Bennett attempted to pose Pteranodon in a once traditional configuration with a forward-facing hand: it didn't go well (note that the walking fingers are even upside down!). |
And there are other issues, too. I wonder why the wing casts exercise was reconstructed with the wing finger joint facing posteriorly rather than laterally, as we'd expect from other pterodactyloids. If the wing finger joint faces backwards the wing spar is almost certainly going to tuck under the arm because the elbow has to bow out from the shoulder when a pterosaur stands: this seems like a foregone conclusion of positioning the wing elements to me, not an unexpected finding. Also of relevance here is that the Q. lawsoni wing metacarpals are, reportedly, poorly-preserved at the proximal (wrist) ends: as with our discussion of the hip bones, above, I wonder if the material is well-enough preserved to substantiate such bold claims? And what of Zhejiangopterus, with its well-behaved wing fingers? Why, again, is Q. lawsoni so different to other azhdarchoids?
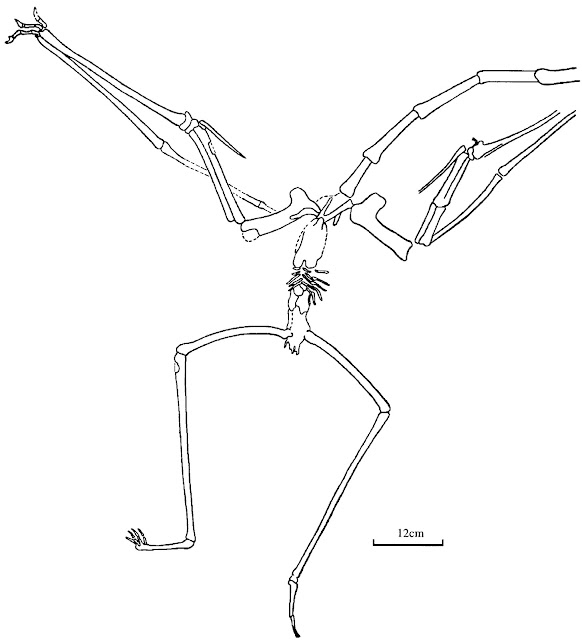 |
| Zhejiangopterus linhaiensis as illustrated by Cai and Wei (1994). OK, this is hardly the height of palaeontological visualisation (to be fair, the original fossil is barely more than an outline) but you can see lots of important functional features in this articulated azhdarchid specimen including splayed hindlimbs, medially-facing palms, and wing fingers that fold up against the outside of the wing. This is all good data that any interpretation of Q. lawsoni functionality needs to be considered against: it's our only direct insight into how azhdarchid skeletons fitted together. |
In sum, I'm not saying that an elbow tucked interpretation is outright wrong. I am, however, very sceptical given the above points and would want to see further research — ideally informed by previous studies on pterosaur wing folding, and bringing in data from Zhejiangopterus and Haenamichnus as well — to substantiate an elbow-tucked wing finger. As you'll note in the art above, I've stuck to convention on this matter with my latest Q. lawsoni piece.
Bipedal launching: back on the table?
Finally, another area of contention between the authors of Padian et al. concerns launch strategy: how did Quetzalcoatlus become airborne? Here, the split is once again between Jim and John on one side, who advocate quadrupedal launch, and Kevin Padian on the other, who prefers a bipedal launch model. This split is not surprising because, some years ago, Jim independently drew the same conclusion about pterosaur launch as Mike Habib, who wrote the first paper on flying reptile quad-launch in 2008. Since then, this idea has become the pterosaur launch hypothesis to beat. As outlined at length in this post, it’s the only concept that explains (following substantial quantification and experimentation) everything we understand about pterosaur size, proportions and muscle volumes, while also fitting launch expectations from pterosaur trackways (i.e. that, among living animals, the gait used for terrestrial locomotion is the same gait used for take-off). It also avoids having to downsize pterosaurs to ridiculously small masses to achieve flight, as exemplified by Chatterjee and Templin’s (2004) conclusion that a giraffe-sized azhdarchid must mass 75 kg or less to facilitate take-off. Under quad-launch theory, giant pterosaurs can easily be 200 or 300 kg and still become airborne (Habib 2008, 2013; Witton and Habib 2010; Habib and Cunningham 2013).
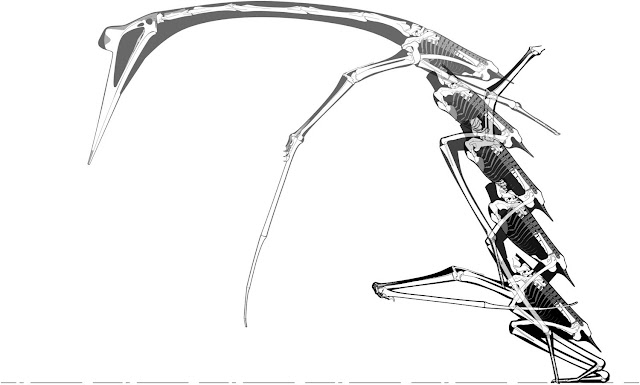 |
| A Q. lawsoni standing-start bipedal launch, illustrated in Padian et al. (2021). Is this a viable launch mechanic for a 4.2 m wingspan pterosaur? Opinion is split among the Padian et al. authorship. |
It’s against this that Padian et al. refloat the idea of Quetzalcoatlus being a bipedal launcher, a concept also emphasised in press releases. The discussions of both bipedal and quadrupedal launch in Padian et al. (2021) are qualitative, mostly focusing on how the skeleton of Quetzalcoatlus can be moved into various launching postures, and there’s little engagement with what’s been said about pterosaur launch by recent workers. For example, Padian et al. suggest that Quetzalcoatlus might have lacked forelimb bending strength to sustain quad launch (i.e. that the forelimb would fail under such stress), ignoring the fact that Mike Habib and I demonstrated over a decade ago that the Q. lawsoni humerus was five times stronger under bending than the femur, and that even the neck bones of Quetzalcoatlus were stronger than its legs (Witton and Habib 2010). There is no explanation for why Quetzalcoatlus lacks the robust hindlimb anatomy of a large hindlimb launcher, when Mike has demonstrated that any flying animal above 500 g starts augmenting its launch limb anatomy to achieve sufficient power and reinforcement to sustain take-off (Habib 2008). And there’s no discussion of why previous calculations of bipedal launch, which have been universally hamstrung by having to lower pterosaur masses to ridiculous levels, went wrong. Padian et al. prefer a (relatively low) mass of 150 kg for a giant azhdarchid, but even this would be two- or three-times too heavy for any published bipedal launch mechanic. The fact bipedal launch proponents have consistently failed to get realistically-massed pterosaurs airborne isn't something we can just ignore: it's evidence against their hypothesis. There’s more we could say, but you're getting the idea: this attempted resurrection of bipedal launch as a viable take-off mechanic for even the small Quetzalcoatlus species has not, in my view, been well-argued, and does nothing to displace quad-launch as the superior pterosaur take-off hypothesis.
Conclusion: extraordinary claims... etc., etc...
And that, I think, is all I want to say on this for now. In short, I can't buy that Quetzalcoatlus is anywhere near as strange as the conclusions of Padian et al. (2021) imply: I strongly suspect it wasn't walking around half-crouched, wasn't flying with its legs tucked up like a bird, and wasn't leaping into the air using its legs alone. But, to be clear, there is nothing wrong with arguing that Quetzalcoatlus was aberrant. However, if you're going to make such assertions you need to present excellent, thorough and fully watertight analyses, and I just don't think Padian et al. do this. There are too many unaddressed complications, overlooked counterarguments and obvious questions raised around their more unusual hypotheses to take them as read. The fact that a lot of the proposals attempt to resurrect somewhat forgotten, decades-old hypotheses should not have been overlooked, either: if we've moved on from those ideas once, why are they suddenly viable now? What's changed to make hip-anchored membranes and bird-like knees plausible for pterosaurs in the 2020s? What are the problems with the consensus views that this paper conflicts with so often? It's this lack of consideration and engagement with modern pterosaur science that is at the core of my scepticism with so much of the paper. But this, of course, is only my take: it’ll be interesting to see what other researchers make of this now that Quetzalcoatlus is finally, and happily, available for unrestricted research access.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored through Patreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusive Patreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
- Andres, B., & Langston Jr, W. (2021). Morphology and taxonomy of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology, 41(sup1), 46-202.
- Averianov, A. O. (2010). The osteology of Azhdarcho lancicollis Nessov, 1984 (Pterosauria, Azhdarchidae) from the late Cretaceous of Uzbekistan. Proceedings of the Zoological Institute RAS, 314(3), 264-317.
- Bennett, S. C. (1990). A pterodactyloid pterosaur pelvis from the Santana Formation of Brazil: implications for terrestrial locomotion. Journal of Vertebrate Paleontology, 10(1), 80-85.
- Bennett, S. C. (1997). Terrestrial locomotion of pterosaurs: a reconstruction based on Pteraichnus trackways. Journal of Vertebrate Paleontology, 17(1), 104-113.
- Bennett, S. C. (2001). The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon Part II. Size and functional morphology. Palaeontographica Abteilung A, 113-153.
- Cai, Z & Wei, F. (1994). On a new pterosaur (Zhejiangopterus linhaiensis gen. et sp. nov.) from Upper Cretaceous in Linhai, Zhejiang, China. Vertebrata PalAsiatica, 32(03), 181.
- Chatterjee, S., & Templin, R. J. (2004). Posture, locomotion, and paleoecology of pterosaurs (Vol. 376). Geological Society of America.
- Costa, F. R., Rocha-Barbosa, O., & Kellner, A. W. A. (2014). A biomechanical approach on the optimal stance of Anhanguera piscator (Pterodactyloidea) and its implications for pterosaur gait on land. Historical Biology, 26(5), 582-590.
- Elgin, R. A., Hone, D. W., & Frey, E. (2011). The extent of the pterosaur flight membrane. Acta Palaeontologica Polonica, 56(1), 99-111.
- Fastnacht, M. (2005). The first dsungaripterid pterosaur from the Kimmeridgian of Germany and the biomechanics of pterosaur long bones. Acta Palaeontologica Polonica, 50(2).
- Frigot, R. A. (2018). Pelvic musculature of Vectidraco daisymorrisae and consequences for pterosaur locomotion. Geological Society, London, Special Publications, 455(1), 45-55.
- Godfrey, S.J., & Currie, P.J. (2005). Pterosaurs. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 292-311.
- Habib, M. B. (2008). Comparative evidence for quadrupedal launch in pterosaurs. Zitteliana, B28, 159-166.
- Habib, M. (2013). Constraining the air giants: limits on size in flying animals as an example of constraint-based biomechanical theories of form. Biological Theory, 8(3), 245-252.
- Habib, M. B., & Cunningham, J. (2010). Capacity for water launch in Anhanguera and Quetzalcoatlus. Acta Geoscientica Sinica, 31, 24-25.
- Kellner, A. W., & Langston Jr, W. (1996). Cranial remains of Quetzalcoatlus (pterosauria, Azhdarchidae) from late cretaceous sediments of big bend national park, Texas. Journal of Vertebrate Paleontology, 16(2), 222-231.
- Hwang, K. G., Huh, M. I. N., Lockley, M. G., Unwin, D. M., & Wright, J. L. (2002). New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri formation, southwestern Korea. Geological Magazine, 139(4), 421-435.
- Lü, J., Unwin, D. M., Xu, L., & Zhang, X. (2008). A new azhdarchoid pterosaur from the Lower Cretaceous of China and its implications for pterosaur phylogeny and evolution. Naturwissenschaften, 95(9), 891-897.
- Mazin, J. M., Billon-Bruyat, J. P., Hantzpergue, P., & Lafaurie, G. (2003). Ichnological evidence for quadrupedal locomotion in pterodactyloid pterosaurs: trackways from the Late Jurassic of Crayssac (southwestern France). Geological Society, London, Special Publications, 217(1), 283-296.
- Naish, D., & Witton, M. P. (2017). Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators. PeerJ, 5, e2908.
- Padian, K. (1983a). Osteology and functional morphology of Dimorphodon macronyx (Buckland) (Pterosauria: Rhamphorhynchoidea) based on new material in the Yale Peabody Museum. Postilla, 189, 1-44.
- Padian, K. (1983b). A functional analysis of flying and walking in pterosaurs. Paleobiology, 9(3), 218-239.
- Padian, K. (1988). The flight of pterosaurs. Natural History, 12, 58-65.
- Padian, K. (2003). Pterosaur stance and gait and the interpretation of trackways. Ichnos, 10(2-4), 115-126.
- Padian, K. (2008). Were pterosaur ancestors bipedal or quadrupedal?: morphometric, functional, and phylogenetic considerations. Zitteliana, B28, 21-33.
- Padian, K., Cunningham, J. R., Langston Jr, W., & Conway, J. (2021). Functional morphology of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology, 41(sup1), 218-251.
- Padian, K., & Olsen, P. E. (1984). The fossil trackway Pteraichnus: not pterosaurian, but crocodilian. Journal of Paleontology, 178-184.
- Paul, G. S. (2002). Dinosaurs of the air: the evolution and loss of flight in dinosaurs and birds. JHU Press.
- Steel, L., Martill, D. M., Kirk, J. R. J., Anders, A., Loveridge, R. F., Frey, E., & Martin, J. G. (1997). Arambourgiania philadelphiae: giant wings in small halls. The Geological Curator, 6, 305-313.
- Unwin, D. M. (1996). Pterosaur tracks and the terrestrial ability of pterosaurs. Lethaia, 29(4), 373-386.
- Unwin, D. M., Lü, J., & Bakhurina, N. N. (2000). On the systematic and stratigraphic significance of pterosaurs from the Lower Cretaceous Yixian Formation (Jehol Group) of Liaoning, China. Fossil Record, 3(1), 181-206.
- Wellnhofer, P. (1988). Terrestrial locomotion in pterosaurs. Historical biology, 1(1), 3-16.
- Wilkinson, M. T. (2008). Three-dimensional geometry of a pterosaur wing skeleton, and its implications for aerial and terrestrial locomotion. Zoological Journal of the Linnean Society, 154(1), 27-69.
- Witton, M. P. (2013). Pterosaurs: Natural History, Evolution, Anatomy. Princeton University Press.
- Witton, M. P., & Habib, M. B. (2010). On the size and flight diversity of giant pterosaurs, the use of birds as pterosaur analogues and comments on pterosaur flightlessness. PloS one, 5(11), e13982.
- Witton, M. P., & Naish, D. (2008). A reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS one, 3(5), e2271.
- Witton, M. P., & Naish, D. (2013). Azhdarchid pterosaurs: water-trawling pelican mimics or “terrestrial stalkers”?. Acta Palaeontologica Polonica, 60(3), 651-660.
Great essay and excellent rebuttal, Mark.
ReplyDeleteThe second I saw Padian's name on the paper, I knew we were in for a trip through the Wayback Machine. Now, it bothers me--it really does--when people just ignore decades of good, reliable data (or throw it all out the window) and insert their long-held pet theories instead. It's intellectually dishonest, especially when those pet theory conclusions cannot be drawn from the material being studied, as is the case here. I find it disrespectful and disappointing.
I do not know Padian personally, but from what I have read here, it definitely sounds like he had his mind made up already and interpreted the evidence based on his preconceptions, rather than letting the data guide him. I could be completely wrong, as he may know of things that I am not privy to. At the least, I hope this controversial paper spurs further research and debate.
DeleteIf Padian is basing his view in the paper on things you are not privy to, then he should have put them in the paper or held off until he could also publish those things. Scientific papers shouldn't rely on folks' willingness to buy a pig in a poke.
DeleteGreat rebuttal, Padian's paper really does seem like a blast from the past in the worst way imaginable
ReplyDeleteShame he can't let go of bipedal pterosaurs, he is otherwise a decent researcher.
ReplyDeleteI also disagree with his assessment of Quetzalcoatlus being a wader, when most evidence suggests azhdarchids were fully terrestrial and lacked adaptations for wading in soft substrates. However, the concentration of fossils in saline lakes is sus.
Well, Mark, that was not nearly as dry as you made it out to be. Measured, thorough and fair. It would have been juicy if you'd just laid into the paper like I do people who are wrong on the internet, but that's not you.
ReplyDeleteI think you make a very good case that the support for the oddness of Quetz among pterosaurs and even azhdarchids is very weak indeed. That there was no biomechanics to the bipedal launch, when the quad launch is supported by such data and calculation makes me think that went like a guy going, 'Hey, what if they ...?' to which someone replies 'You should look into that.' but it went no further. Near as I can make out, the best that can be said for Padian's ideas on how Quetz fit together is 'It's not impossible it was like this.'
And I have to play language pedant and note that you used "credulous" = 'having or showing too great a readiness to believe things.', which seems to be Padian when it comes to seeing things as birds, where you should have used "credible" = 'believable' or 'convincing'. That is a hill I am prepared to die on!
BTW, having just said that, I wonder if Padian's early work on birds as dinosaurs didn't predispose him to see Quetz as more birdlike than it had any reason to be.
Thanks for the typo spot! I'm also curious as to how some of the arguments in this paper came into being as it certainly is - with the frequent author disagreements and all - a strange publication. It can't have helped to have had two sets of authors disagreeing on basic, fundamental aspects of biomechanics. Perhaps the efforts to compromise led to awkward fence-sitting from both parties, rather than more substantial investigations from either side?
DeleteNit-pick: "It really needs bolstering with more data to be credulous". That should be "credible", meaning "capable of being believed". "Credulous" means "Having a tendency to believe things too easily".
ReplyDeleteThat bit of pedantry aside (which I now see the mysterious Michael had already dealt with), what an amazing post this is. By far the most polite and measured atomic smackdown I've ever read. Mark, I don't know whether to congratulate you for your technical insights or for your equanimity.
ReplyDeleteThanks for spotting that typo, Mike, and for the nice words about the post. If this reads as especially polite and considerate, it's only because I sometimes wonder if I've been a little mean in the past. As much as I want to share my takes on certain topics, I also don't want to be insulting or tick anyone off. There's enough people doing that elsewhere online!
DeleteIs it possible that much of this paper was actually written in the 80s, but only published now?
ReplyDelete"ignoring the fact that Mike Habib and I demonstrated over a decade ago that the Q. lawsoni humerus was five times stronger under bending than the femur, and that even the neck bones of Quetzalcoatlus were stronger than its legs"
ReplyDeleteCan't wait for neck-launching pterosaurs. Although Hatz would probably be better at that than Quetz.
Michael Bech, I bet you're right.
ReplyDeletePds3.14, LOL at neck-launching!
So out of curiosity, what happens in cases where you have a team of researchers working on a publication and they favor differing interpretations of their own work? I'm just wondering how you get to a situation like this paper where 2/3 of the authors disagree with some of the main conclusions of the study itself and its lead author?
ReplyDelete