
 |
| The 2015 version of the giraffe vs. azhdarchid vs. person image, now in it's fifth iteration (see the general history of these images through the years: 2006; 2007; 2009). The giraffe is a big bull Masai individual, standing a healthy 5.6 m tall, close to the maximum known Masai giraffe height. The pterosaur is a 10 m wingspan Arambourgiania philadelphiae (for reasons I cannot go into now, it is not wise to consider the appearance of giant azhdarchid taxa interchangeable any more: this should not be considered Hatzegopteryx thambema or Quetzalcoatlus northropi). The Disaknowlegement provides the human touch. These characters will receive some additional company soon. |
They nearly weren't called 'azhdarchids'
'Azhdarchidae' is a terrific name. It's short but mysterious, relatively easy to spell, and PR friendly enough that even the British tabloid The Sun has used the term on at least two (I think) occasions. The name 'Azhdarchinae' was coined by the late Lev Alexandrovich Nesov in 1984 from the Uzbek word 'azhdarkho', a name for a mythical dragon, and also the nomenclatural basis for the medium-sized Uzbek azhdarchid Azhdarcho lancicollis. Nesov's name encompassed all three azhdarchid genera known at that time: Azhdarcho, Titanopteryx (now known as Armabourgiania) and Quetzalcoatlus. Almost simultaneously, however, the exact same set of taxa was being roped into another group by Kevin Padian, which he termed Titanopterygiidae after, obviously, Titanopteryx. Nesov's 'Azhdarchinae' pipped the far-less elegant Titanopterygiidae to the publishing punch by a matter of months, and took nomenclatural priority for the group. Padian elevated Azhdarchinae to 'family' level in a short note in 1986, giving us our now familiar term, 'Azhdarchidae'.
 |
| Lev Alexandrovich Nesov holds the fossil cervical vertebra, notarium and jaw tip of the azhdarchid Azhdarcho. Image from Unwin (2005). |
Despite their giraffian proportions, giant azhdarchid torso were relatively tiny. Witton and Habib (2010) noted that, like many pterodactyloid pterosaurs, their torsos were probably only a third or so longer than their humeri, suggesting a shoulder-hip length of about 65-75 cm for an animal with a 10 m wingspan. That's a torso length not much larger than your own, although they were considerably more stocky and swamped with muscle. Azhdarchid shoulders, in particular, are well endowed with attachment sites for flight muscles, as are (for pterosaurs) their pelves and hindquarters.
Giant azhdarchids did not suffer from flight power shortages
Many internet commenters often roll out the idea that giant azhdarchids would struggle to take off from the ground, even allowing for new ideas like quadrupedal launching. These folks need to get out of their armchairs, however, and check out some classic work on animal flight and giant pterosaur takeoff. James Marden's 1994 work on animal takeoff found some surprisingly consistent scaling trends among animal flight power and takeoff ability, allowing us to predict the muscle power of even long extinct fliers like Meganeura, Archaeopteryx and a 10 m span azhdarchid. The resulting aerobic power output of azhdarchid flight muscles - all 60 kg of them (a fairly safe bet for a 250 kg azhdarchid given what we know of animal flight muscle fractions among modern fliers) - is a bit rubbish, only 4.52 N/kg of body weight. Animals need to be generating 9.8 N/kg to fight gravity, so this would seemingly ground our giants. Bear in mind, however, that swans, albatross, vultures and turkeys also have aerobic power outputs of around 4.5 N/kg from their flight muscles, and they can fly just fine. The secret to their takeoff lies in the great power of anaerobic muscle contraction, which provides twice the power achieved under aerobic regimes. Using anaerobic power, giant azhdarchid power outputs are 10.098 N/kg of body weight, a value surpassing the 9.8 N/kg and matching the anaerobic power outputs of a 10 kg swan or 1 kg vulture (see graph, below). In terms of power availability, then, giant azhdarchids would not have struggled to launch any more than a large bird, so all these suggestions about poor takeoff ability and whatnot can be put to bed.
 |
| Scaling of flight performance with body size under anaerobic power output. The dashed line is the minimum lift needed to overcome gravity. Anaerobic power is 225W/kg, the upper limit of avian anaerobic output. From Marden (1994). |
Azhdarchids are undeniably best known from Upper Cretaceous rocks, but they also have a patchy and sometimes controversial Lower Cretaceous record. Recently, Gareth Dyke and colleagues (2011) demonstrated that the group were probably present at the very base of the Cretaceous, in Berriasian (c. 140-145 Ma) deposits of Romania. Given that azhdarchids are definitely present at the final stage of the Cretaceous, this gives the group a stratigraphic record spanning the entire Cretaceous: 80 million years in total. This is longer than any other pterosaur group. Two cervical vertebrae from the Late Jurassic of Africa may extend their temporal range another 5 million years, although the affinity of these specimens remains controversial.
 |
| A much improved skeletal reconstruction of the small azhdarchid Zhejiangotperus linhaeiensis over my oft-reproduced effort from Witton and Naish 2008. Note the use of pacing strides, a gait indicated by pterosaur trackways but seldom seen in pterosaur palaeoart. |
When we describe azhdarchids, we often use two qualifiers: 'toothless' and 'long-necked'. In fact, these pterosaurs are brimming with characterising features (above). Their rostra are particularly elongate compared to all other pterosaurs, their orbits are depressed well into the lower half of their skulls, their wing metacarpals and femora are atypically long, and their extremities are short and robust. Their mid-series cervical vertebrae are famously simplified into almost tube-like structures, and their humeri are deceptively derived from the pterodactyloid norm. The wing fingers of azhdarchids occupy a relatively small 47% of their wing lengths, a value only approximated by one other pterosaur group, the closely related thalassodromids. Artists, take note: grounded azhdarchids should not be reconstructed with their folded wing fingers stretching skywards over their backs: they couldn't reach that far.
But no, seriously, the long necks
The cervical vertebrae of giant azhdarchids are poorly known, with only a few specimens (and even fewer good ones) being recovered to date. These rare fossils do, however, clearly indicate substantial neck proportions in at least animals like Arambourgiania. The holotype cervical V of this animal is around 660 mm long, and is missing an estimated 100 mm from its posterior end. Steel et al. (1997) scaled this vertebra isometrically with relatively complete neck skeleton material from the 4.7 m wingspan azhdarchid Quetzalcoatlus sp. to predict a whopping 3071 mm length for cervicals III-IX in Arambourgiania. The use of isometry here is questionable (Witton and Habib 2010), but is defensible given the amount of azhdarchid neck material available to these authors in the mid-nineties. Ongoing work I'm involved with (which will hopefully be published before we're too much older) has attempted to apply allometry to calculations of giant azhdarchid neck lengths. The results are a little more conservative than the 3 m offered here, but we're still landing in the "seriously long neck" camp. Whether azhdarchids will retain the title of absolutely longest necks outside of Sauropoda (Taylor and Wedel 2013) remains to be seen however: I suspect they may ultimately just be pipped by the weirdo protorosaur Tanystropheus. Dammit.
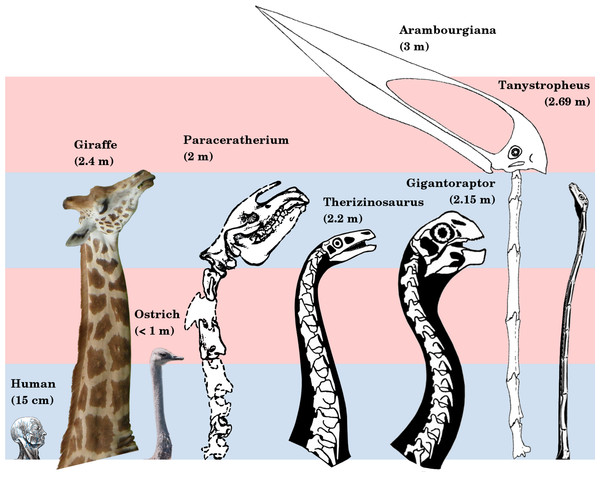 |
| The 'Big Necks Which Don't Belong to Sauropods Competition', won by the giant azhdarchid Arambourgiania. From Taylor and Wedel (2013). |
The necks of azhdarchids are not just famous for their size, but are also renowned for their rather inflexible joints. These widely discussed features have been the bane of many azhdarchid lifestyle interpretations (see Witton and Naish 2008 for a review), but actual quantification of their arthrological range has been lacking until recently. This is, in part, because a complete 3D azhdarchid cervical series has been elusive for many years, but Alex Averianov (2013) recently produced a composite digital neck skeleton for Azhdarcho to figure out their range of motion. The results were more-or-less what we all expected: very limited range in the mid-series, with most of the mobility limited to the extremes. A surprising amount (but still fairly restricted) range of motion was afforded at the neck base, however. As may be expected, this study is very welcome to those of us interested in the biomechanics and functional anatomy of these animals, and I'm glad to see it.
 |
| Averianov's (2013) reconstructed neck arthrology of Azhdarcho lancicollis. That's one stiff neck. |
Swimming piscivores and aerial hawking: genuinely suggested azhdarchid lifestyles
It's well known that most recent 'serious' proposals of azhdarchid lifestyles are things like skim-feeding, terrestrial stalking and wading, but many other, frankly outlandish palaeoecological hypotheses have been thrown at azhdarchids over the years. Lev Nesov perhaps takes home the prize for the most bizarre ideas, proposing in his 1984 paper that azhdarchids could swim to find food (both along the surface and by diving) and pursue 'poorly flying' vertebrates through the air. In the same paper, he also advocates skim-feeding as a probably azhdarchid lifestyle. I remain unsure which part of azhdarchid anatomy indicated to Nesov that these animals had superhero-like abilities to acquire food.
 |
| Sauropods give a giant azhdarchid the evils. Seems they don't like being buzzed at close range. |
Although azhdarchids are frequently discussed for their natty terrestrial capability nowadays, it's important to remember than any substantial travelling they had to do was probably performed in the air. Computations of the flight abilities of giant azhdarchids have returned seriously impressive results (Witton and Habib 2010). As mentioned above, azhdarchids likely employed anaerobic power for strenuous flight activities like takeoff and perhaps flapping bursts, and likely relied mostly on thermal soaring and flap-gliding like modern raptors to remain airborne for long periods. Their minimum sink and best glide speeds are steady cruises at 16.3 - 24.9 m/s (58.7 - 89.4 kph) but, if they were in a hurry (such as looking for a source of uplift), speeds of up to 48.3m/s (173 kph) were possible for short durations. We estimated that azhdarchids had about 90 - 120 seconds of anaerobic burst power before tiring, meaning these animals could go from a standing start to - literally - several kilometres away in the space of a few minutes. Yowsers. What's more, the size and bodily resources available to such large creatures permitted tremendous flight times: up to 16,000 km of travelling without resting or foraging were likely possible. That's the equivalent of an animal flying from London to Vegas non-stop, realising it forgot its passport, and then flying home again without touching the ground.
And that's your lot for now, folks. If you want to know more about azhdarchids, be sure to check out my book for a whole chapter about them, which is something like the second biggest entry in the entire thing. Things may go quiet over the next few weeks while I'm away at various conferences, but posting will resume when I get back.
References
- Averianov, A. O. (2013). Reconstruction of the neck of Azhdarcho lancicollis and lifestyle of azhdarchids (Pterosauria, Azhdarchidae). Paleontological Journal, 47(2), 203-209.
- Dyke, G. J., Benton, M. J., Posmosanu, E., & Naish, D. (2011). Early Cretaceous (Berriasian) birds and pterosaurs from the Cornet bauxite mine, Romania. Palaeontology, 54(1), 79-95.
- Marden, J. H. (1994). From damselflies to pterosaurs: how burst and sustainable flight performance scale with size. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 266(4), R1077-R1084.
- Nesov, L. A. (1984). Pterosaurs and birds of the Late Cretaceous of Central Asia. Paläontologische Zeitschrift, 1, 47-57.
- Padian, K. (1984). A large pterodactyloid pterosaur from the Two Medicine Formation (Campanian) of Montana. Journal of Vertebrate Paleontology, 4(4), 516-524.
- Padian, K. (1986). A taxonomic note on two pterodactyloid families. Journal of Vertebrate Paleontology, 6(3), 289-289.
- Steel, L., Martill, D. M., Kirk, J. R. J., Anders, A., Loveridge, R. F., Frey, E. & Martin, J. G. (1997). Arambourgiania philidelphiae: giant wings in small halls. The Geological Curator, 6, 305-313.
- Taylor, M. P., & Wedel, M. J. (2013). Why sauropods had long necks; and why giraffes have short necks. PeerJ, 1, e36.
- Unwin, D. M. (2005). The pterosaurs from deep time. Pi Press, New York.
- Witton, M. P., & Habib, M. B. (2010). On the size and flight diversity of giant pterosaurs, the use of birds as pterosaur analogues and comments on pterosaur flightlessness. PloS one, 5(11), e13982.
- Witton, M. P., & Naish, D. (2008). A reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS One, 3(5), e2271.
"[T]his gives the group a stratigraphic record spanning the entire Cretaceous: 80 million years in total. This is longer than any other pterosaur group."
ReplyDeleteBut, given that they must have originated well after the ancestral pterodactyloid, wouldn't the other branch of Pterodactyloidea (Ornithocheiroidea, I think) be about as long, even if they snuffed it earlier?
If ghost lineages rather than stratigraphic occurrences are taken into account, nyctosaurs (members of Ornithocheiroidea according to some) may match azhdarchids, but only in some interpretations of pterodactyloid phylogeny. My preferred interpretation of the pterosaur tree (something more-or-less like the recent variants of the 'Unwin phylogeny') wouldn't show this. The longest lineage of ornithocheiroids - Ornithocheiridae itself - are probably the current runner up for the longest-lived pterosaur lineage, at 60 Ma.
DeleteAbout about the New Zealand pterosaur?
Delete"The only pterosaur fossils described from New Zealand are the
distal extremity of a left ulna and a partial tooth, collected from
the Late Cretaceous (Campanian–Maastrichtian) Mata series at
Mangahouanga Stream on the North Island (Wiffen and Molnar,
1988; Molnar and Wiffen, 1994). The identity of this specimen is
also uncertain, although Wiffen and Molnar (1988) reported that
the ulna was most similar to Anhanguera araripensis (sensu Kellner and Tomida, 2000) and Dsungaripterus weii (sensu Young,1964)."
If it's an ornithocheirid, it would make Ornithocheiridae as long lived as Azhdarchidae (alongside Piksi?). If it is a dsungaripteroid, then dsungaripteroids are the unambiguously longest lived pterosaur clade known, adding an extra 30 million years.
The New Zealand specimen (a partial ulna and tooth, if memory serves) is pretty undiagnostic, other than being belonging to a reasonably sized pterodactyloid. It doesn't really help us establish the ranges of anything. Note that isolated 'pterosaur' teeth are often misidentified titanosaur or croc teeth, too.
DeleteThere's need to be careful with definitions of Dsungaripteroidea, too. There's a bit of a slop about the use of that name, referring to either a Germanodactylus + dsungaripterid group in some phylogenies (which has a stratigraphic range of Late Jurassic - mid-Cretaceous) or a much broader clade of pterodactyloids (essentially ornithocherioids, azhdarchoids and dsungaripterids). The latter is certainly a long lived group, but represents a clade of pterosaurs well above the 'family' level we're working at here.
As for Piksi, I'm not 100% convinced it represents an ornithocheiroid.
I thought that Dsungaripteroidea as "all notarium based pterosaurs" was only used once? At any rate, I think the context was on the Germanodactylus + Dsungaripteridae context.
DeleteYoung (1964) defined Dsungaripteroidea as, in essence, a clade comprised of notarium-bearing pterosaurs. The definition has since been tweaked by several different authors to mean quite different things, so any mention of 'Dsungaripteroidea' needs a qualifier to make it clear which definition we're following.
DeleteThis comment has been removed by a blog administrator.
DeleteNesov's 'Azhdarchinae' pipped the far-less elegant Titanopterygidae to the publishing punch by a matter of months, and took nomenclatural priority for the group.
ReplyDeleteThat actually doesn't matter. The reason why Arambourgiania isn't called Titanopteryx anymore is that the name T. was preoccupied by an insect. Any family Titanopterygidae must be based on the valid name T., not on its invalid junior homonym.
I think there's been some misunderstanding here. I wasn't referring to the replacement name for Arambourgiania (I'm very familiar with that story), but the naming of the azhdarchid clade itself. Nesov's publication specifying the content, name and diagnosis for the group beat Padian's similar work by a mere few months and, under ICZN Guidelines, had precedent. My point is that the group missed out on having a pretty horrible name by a relative whisker, not anything about the validity of Titanopteryx as the basis of a pterosaur clade name (which, of course, you're correct about).
DeleteThis is irrelevant. Titanopterygidae follows Titanopteryx, which based on preoccupation is an insect. This means that Titanopterygidae is not merely incorrectly formed, it must be shifted as a "family" rank taxon for the insect clade. It doesn't compete for priority with Azhdarchidae under any circumstance because the two names do not have concordant contents. Moreover, only under assumptions that the species taxa Azhdarcho and Arambourgiana(Titanopteryx) were synonyms would the case that Titanopterygidae ever compete directly with Azhdarchidae (under the Principle of Coordination). But, because they also lack the same type species, the only type of synonymy that would be proposed would be the subjective kind, where people play games about whether Thalassodromidae should be in Tapejaridae if they were sister taxa (which I am not arguing for or against at this moment).
DeleteI'm not sure I agree that this case is particularly subjective. The formulation of the two proposed 'families' (they weren't viewed as clades at the time, of course) is surely similar enough that, sidestepping the preoccupation issue of Titanopteryx, we had almost simultaneous competing labels for the clan we now know as azhdarchids. Even Padian himself said so in his 1986 paper, which was written specifically to address this issue. In his introduction, he states:
Delete"Shortly before my diagnosis of the new Family Titanopterygiidae was published (Padian, 1984), a paper by L. A. Nesov (1984) appeared in which he named a virtually identical taxon at the sub-familial level. This brief note is to emend the taxonomy and revise the diagnoses of the taxa involved."
And in later elaboration:
"Nesov (1984) named the Sub-family Azhdarchinae of the family Pteranodontidae on the basis of his new genus and species Azhdarcho lancicollis, from the Upper Turonian-Coniacian of the Uzbek SSR. Nesov's diagnosis leaves no doubt that the characters of the cervical vertebrae, on which I based my diagnosis of Titanopterygiidae, are identical, and therefore his name should take priority."
Setting the confusion of the Arambourgiania/Titanopteryx to one side for the moment, my point is that Padian's proposed name for the azhdarchid group almost won out, at least temporarily. I don't think workers of that time were in any doubt that he and Nesov had identified the same basic group, and only one name would have stood. 'Azhdarchidae' would probably have been considered an acceptable replacement once the synonymy of Titanopteryx was discovered, of course.
Thanks Mark. Always edifying.
ReplyDeleteThis comment has been removed by the author.
ReplyDeleteI have heard somewhere that a certain Eurasian giant azhdarchid (the largest, the island giant-you know which one I mean) was very robustly built for hunting l=big prey, is this valid?
ReplyDeleteMy understanding is that Hatzegopteryx was indeed adept at eating relatively heavy prey. It may well have been capable of eating prey up to the size of a human whole, and it could almost certainly have been highly effective as a storker.
ReplyDeleteMy understanding is that Hatzegopteryx was indeed adept at eating relatively heavy prey. It may well have been capable of eating prey up to the size of a human whole, and it could almost certainly have been highly effective as a storker.
ReplyDeleteVerry awesome! I love your articles! BTW, have you ever thought about creating a DeviantArt account? The paleo community misses you there!
ReplyDeleteVerry awesome! I love your articles! BTW, have you ever thought about creating a DeviantArt account? The paleo community misses you there!
ReplyDeleteWould it be fair to say that so far no (zero) large mature pterosaurs exhibit owl-like eyes forward binocular vision? Is there any indication that eye placement might change from chick to eyes on the sides adult as the skull (and perhaps crest) grew and eventually fused? The questions were actually inspired by thinking about how in The Lord of the Rings Eowyn (on foot)
ReplyDeletefights a fell beast that the author agreed resembled a pterodactyl (although that was not his inspiration). I would not fancy the odds of her severing the neck from back to front with a one-handed sword (technically, probably a saber) – any thoughts? The fell beast would probably be something the dimensions of an azhdarchid - peterfzoll@yahoo.com
This comment has been removed by a blog administrator.
ReplyDelete